| PART/CELL NAME |
ABBREVIATION
SYNONYMS (S)
ANTONYMS (A) |
LINEAGE |
DESCRIPTION |
G1
G2 |
|
ABprpaaaapa
ABplapaapa |
Ventral ganglion neuroblasts present at hatching, located near the excretory pore. G1 divides in L1 to produce ring motor neurons/ interneurons RMHR (G1.r) and RMHL (G1.l ). G2 divides in L2 to ring motor neurons /interneurons RMFR (G2.ar) and RMFL (G2.al) and excretory socket/pore cell (G2.p).
See Excretory
system
|
| G
actin |
Globular actin (S)
Monomeric actin (S) |
|
G actin is the monomeric form of actin that is often stored in large pools so that it is available to form long polymers in the presence of ATP. These polymers are called F actin (filamentous actin). G actin can also assemble with certain actin-binding proteins to form a gel-like
network, a common cytoskeletal feature found in the lamellipodia of migrating
cells or as reinforcement to the plasma membrane in cells under stress.
See Actin
See F actin
|
| “g”
neuron |
ASG cell (S) |
|
See ASG cell |
| Galvanotaxis |
|
|
A behavior in which the animal
orients its body motion to the direction of a local electrical field.
Most nematodes seem to have this capability, moving towards the cathode,
but the nature of the response is not understood. C. elegans has been
observed to move directly towards or away from a current source in an
electric field (Sukul
and Croll, 1978).
See Taxis
|
| Gamete |
|
|
A cell
in the germline that will give rise to one or more germ cells (spermatozoa
or oocytes) by later cell divisions and/or maturation.
Alternately, a gamete or a “mature gamete” can refer to a mature
oocyte or spermatozoon, with a haploid set of chromosomes, that is a cell
that is competent to fuse with another gamete to create a zygote. In C.
elegans, the two precursors of germline tissue are Z2, Z3, the daughters
of embryonic founder cell P4. |
| Gametogenesis |
|
|
The process
through which gametes, ova or spermatozoa, are created.
For greater detail see Germ line
See Oogenesis
See Spermatogenesis |
| Ganglion |
|
|
A cluster of neuron cell bodies (somata). In higher animals, virtually all ganglia also include a local zone of neuropil (synaptic processes), often as a distinct profusion of short branches surrounded by a ring of cell bodies. Thin glial sheets often separate the somata from the neuropil. However, in C. elegans most synapses are centralized in the nerve ring and the major motor nerve cords, so that many ganglia either lack neuropil, or exist as separate cell clusters in loose association with the nerve ring neuropil. Ganglia associated with the nerve ring include the dorsal, lateral (bilateral pair) and retrovesicular. Some ganglia in the tail have local neuropil only in the adult male (lumbar and dorso-rectal). Ganglia associated with the ventral nerve cord include the ventral and pre-anal. |
| Gap junction |
Electrical synapse
(S) |
|
A common (~600 per animal, White et al., 1986) intercellular junction found between opposing plasma membranes of two neighboring cells that allows the intercellular transfer of electrons and small molecules. These generally occur between two like cells within a tissue, but can also occur between nerve and muscle, or between somatic gonad and germline. While in higher vertebrates all gap junctions are formed by members of the connexin gene family, in nematodes and other invertebrates, these junctions appear to be formed only by members of the innexin family (Phelan and Starich, 2001). These junctions may also be sites of metabolic coupling. |
| Gastrula |
|
|
The form of the embryo at the end of gastrulation, at which time the germ layers have moved inside the embryo; the preceding stage is often called the blastula.
|
| Gastrulation |
|
|
A key step in the early development of the embryo, in which some cells from the outside of an relatively undifferentiated, single mass of about 26 cells, begin to migrate inward from the ventral side to create an inner cell layer from which mesodermal tissues will eventually be derived. This step begins about 100 minutes after fertilization and is completed about 200 minutes later. By the end of gastrulation, many cells among both the inside and outside layers have begun to differentiate into distinctive cell types and tissues, and the embryo begins to elongate. The first cells to ingress at gastrulation are the founder cells Ea and Ep (which found the intestine), P4 (which founds the germline), the MS daughters, and some cells stemming from C, D and AB founder cells.
Gastrulation has been described in detail for C. elegans by Sulston et al. (1983), Lee and Goldstein (2003) and by Nance et al. (2005).
See Elongation
|
| Genitalia |
|
|
This term is used to refer to the secondary sexual features of the hermaphrodite and male. Male genitalia includes the male tail, such as the fan, rays, spicules, and gubernaculum while the hermaphrodite's primary feature is the vulva. |
| Genital papilla/papillae |
|
|
This term may sometimes be used to refer collectively to the male tail rays, the hook, spicules, and post-cloacal sensilla, and in older nematode literature was meant to refer specifically to the rays. The term could also refer more literally to an intermediate stage during ray formation, just prior to ray extension, when each ray cell group actually does take the form of a small bump in the cuticle, e.g. a papilla, before the ray extends to adopt its adult appearance. |
| Genital sense organs |
|
|
Refers to the sensilla of the male tail, including those of the spicules, rays, and those surrounding the cloacal opening. |
| Genital tract |
|
|
See Reproductive tract |
| Genome |
|
|
The complete collection of genes encoded within the nucleus of a cell, including both expressed and unexpressed gene sequences. The 97-megabase genomic sequence of C. elegans revealed over 19,000 genes, ~40% of which have predicted protein products that match those in other organisms (C. elegans Sequencing Consortium, 1998). See Piano et al. 2006 for more detail. WormBase is an online resource for information on the genomics of C. elegans. |
| Gentle touch |
Light touch (S) |
|
A mechanosensory behavior, mediated by six touch receptor neurons. Response to gentle touch is tested by stroking the worm with a eyebrow hair (see video). Stroking the anterior portion of the animal results in backward movement while touching the tail causes the animal to move forward (For review see Bounoutas and Chalfie, 2007).
See Harsh touch
See Mechanoreceptor neurons
See Mechanosensation
See Touch response
|
| Geotaxis |
|
|
Directed movement in response to the gravitational field that causes the animal to become oriented to the source. Positive geotaxis is down toward the direction of gravity and carries the animal into the soil, whereas negative geotaxis carries the animal upwards. No response to gravity has yet been demonstrated in C. elegans.
See Taxis
|
| Germ cell |
|
|
Any member cell of the male or female germline, including sperm, oocytes, and their precursors. In the nematode gonad, the germ line proliferates from one primordial germ cell (PGC). Immature germ cells are not completely cellularized; instead they each maintain a syncytial linkage via the rachis, yet also retain their individual plasma membrane borders. Individual “cells” seem to act independently within this syncytium, therefore each connecting element, containing one nucleus, a few cytoplasmic organelles and a bounding plasma membrane is considered a separate germ cell.
See Oocyte
See Spermatocyte
See Rachis
|
| Germ layer |
Embryonic germ layer (S) |
|
In higher animals, this term refers to three principal groups of cells that lie in separate layers and give rise to separate tissues: ectoderm, mesoderm and endoderm. In C. elegans, such cell groups begin as just small cell clusters in the early embryo, rather than as clearly separate layers (ectoderm = AB + C blast cells; mesoderm = MS + C; endoderm = E). Their lineages are not as restricted as expected, so that the AB lineage provides a few mesodermal cells, while the MS lineage provides a few ectodermal cells. Nonetheless, the term is occasionally used within C. elegans literature to reference these few blast cells (von Ehrenstein and Schierenberg, 1980). |
Germ plasm
|
Germinal plasm (S)
Nuage (S) |
|
Cytoplasm found in germline cells but not somatic cells. Germ plasm is composed of specific cytoplasmic components (proteins, P granules, and nuclear DNA) present in the early embryo that segregate only to the progenitor cells of the germline and not to the progenitors of the somatic lineages. This segregation process begins with the very earliest cell divisions in the embryo (Gönczy and Rose, 2005) and restricts germ plasm to the posterior P blastomeres, both by intracellular transport and anchorage of these elements to the posterior pole (Guo and Kemphues, 1995; Boyd et al., 1996; Lyczak et al., 2002), and by active degradative mechanisms (Spike and Strome, 2003; Pellettieri et al., 2003). This active degradation is one of the mechanisms by which the germ plasm matures into embryonic cytoplasm in all somatic cells, while the germline progenitor cells segregate and retain germ plasm at the posterior pole of the embryo. |
| Germinal cell |
Germline precursor (S)
|
|
The founder cells of the germline, Z2 and Z3 (derived from P4), and their daughters. |
| Germinal nucleus |
|
|
Referring to a nucleus of one of the founder cells of the germline (Z2 or Z3), but not including nuclei that give rise to somatic gonad (Z1 or Z4). |
| Germinal vesicle |
|
|
The nucleus of the primary oocyte at the time of fertilization; it is the source of the female pronucleus prior to pronuclear fusion. It generally lies at the opposite pole from the site of sperm entry, and seems to break down (Germinal Vesicle Break Down -GVBD) shortly after the fertilization event (Nigon, 1949; Nigon and Brun, 1955; McNally and McNally, 2005). |
| Germinal zone |
Mitotic zone (S) |
|
The portion of the gonad in which the germ cells are mitotic. In the hermaphrodite, these are the distal portions of the ovaries which are found in the distal portion of each of the two gonad arms. The male germ line also exhibits distal-proximal polarity with the mitotic cell population located at the most distal end of the gonad in the testis. |
| Germline |
|
|
The lineage that gives rise to all germ cells in an animal, that is the spermatozoa, oocytes and their precursors, through repeated cell divisions stemming from a single founder cell, the P4 blastomere.
See also Hubbard and Greenstein, 2005 |
| Germline founder cell |
P4 blastomere |
|
The germline founder cell that gives rise, by mitotic cell divisions, to all of the germ cells, but does not contribute to the somatic portion of the gonad. |
| Germline proliferation |
|
|
The process by which the one primordial germ cell proliferates to become over a thousand cells in the adult animal. Initial proliferation occurs in the distal mitotic zone. Then, via a carefully regulated pathway, cells move proximally to a transition zone where they enter meiosis and later, gametogenesis. Germline proliferation arrests if larvae enter the dauer stage and resumes once normal development occurs. |
| Germline tumor |
|
|
A mutant phenotype involving germline proliferation, in which the germ cells proliferate in unchecked divisions. In one case, they complete the early stages of meiotic prophase, but then exit pachytene and return to a mitotic cycle. Mutations in gld-1 produce this phenotype in hermaphrodites, but not males (Francis et al., 1995). Other studies have shown that an inappropriate interaction between proximal sheath cells and undifferentiated germ cells can drive the formation of a proximal germline tumor implicating the somatic gonadal sheath in the regulation of germline development (Killian and Hubbard, 2005). |
| GFP |
Green fluorescent protein |
|
A commonly used molecular marker used in transgenic strains to label a gene product, organelle, cell or tissue in live animals. The original gene was derived from jellyfish and gives off a green color under fluorescent illumination (Chalfie et al., 1994). Alternate versions are now available which give other colors such as yellow and cyan (Miller et al., 1999). Using GFP in reporter gene fusions in C. elegans has become a well used tool for visualizing gene expression patterns (Boulin et al., 2006).
See CFP
See YFP
|
| Gland cell |
|
|
A variety of very different cell types which share cytoplasmic features (such as large membrane-bound granules) that suggest a role in secretion, thus termed gland cells. Most, but not all, extend long ducts or cell processes away from the cell body to the site of secretion. They lie in different parts of the bodies, in association with very different tissues, and may open externally or internally. Listed below are the gland cell types of C. elegans. Only the pharyngeal glands seem to have direct innervation (Albertson and Thomson, 1976). Although common in some other nematode species, there are no “caudal” glands in C. elegans, which would secrete material out of the extreme tail tip.
Amphid glands = amphid sheath cells with external opening
Pharyngeal glands = g1 (3 cells, one pair fused) and g2 (2 cells) all open to pharynx lumen (see also Pharynx Atlas)
Excretory gland = fusion of two cells and opens to excretory duct
Rectal gland = composed of 3 (non syncytial) cells, opens to intestinal lumen |
| Glial cell |
|
|
In C. elegans, the sheath cells, socket cells and structural cells bear some resemblance to glia, since they provide thin wrapping structures around neuropil or axons, or produce protected environments for nerve endings (dendrites). They may also provide substrates for outgrowth of growth cones (Wadsworth et al., 1996).
See Neuronal support cells.
While the GLR cells have been named as if they might serve a glial function in the nervous system, in fact they appear to belong more properly to the mesoderm and act as scaffold cells to the head muscles, not directly touching the nerve ring itself, and therefore not serving a glial function per se.
See Scaffold cell
|
| Glottis |
Metastom (S) |
|
The portion of the posterior buccal cavity where cuticular flaps operate to open and close the pharyngeal lumen.
See Flaps
|
| Glycocalyx |
Surface coat (S)
Envelope (S) |
|
This term has multiple uses:
1) An outer coating of the cuticle, often called the “surface coat” of the epicuticle. While its molecular makeup is not completely understood, it may include glycoproteins (proteoglycans and mucins) and many tissues may contribute components to it though it is known that it is non-collagenous. There are suggestions that some portions are secreted from the pharyngeal and excretory glands (Singh and Sulston, 1978; Bird and Bird, 1991), the excretory system (Nelson and Riddle, 1984; Bird et al., 1988; and by analogy to T. canis data Blaxter et al., 1992), the sensory sheath cells of the sensilla (Anne Hart’s data), possibly from the lumenal surface of the intestine, material deposited by transcuticular mechanisms from beneath the cuticle (Blaxter and Bird, 1997), or even material secreted by the newly fertilized embryo (Hall, unpublished). Some species of nematodes have specialized glands in the head or tail (epidermal glands) that contribute secretions to the glycocalyx (Bird and Bird, 1991). By analogy, it seems possible that the rectal glands in C. elegans tail may also contribute to the cuticle's glycocalyx. The glycocalyx may serve multiple functions, including as a lubricant (to aid in the animal’s movements through the environment), as a protective agent or in pheromone signaling to other nematodes.
2) The extracellular coating on microvilli of intestinal cells facing the lumen. The molecular nature of this feature has not yet been determined in C. elegans.
3) The extracellular coating over the lumenal surface of the excretory canal sinus. Known to include LET-653 mucin (Guerardel et al., 2001).
4) The extracellular coat on the surface of the uterine epithelium, facing the lumen which may help to cement the lumen into an “open” position to create the egg chamber in the adult gonad (White, Southgate and Hall, unpublished).
5) A covering of the surface of the proximal 2-3 oocytes between the sheath and the oocytes.
6) The temporary coat on the young embryo immediately after fertilization, which forms within seconds and is then shed upon secretion of the eggshell. |
| Glycogen |
|
|
A polysaccharide molecule used as a means to store glucose in certain tissues. It is prominently seen by TEM in muscle arms and in hypodermis in well fed C. elegans adults. These small storage granules have an electron lucent, particulate appearance by TEM (about 10-40 nm diameter), often appearing brighter than any other object, as if they fail to stain or even to be infiltrated by the plastic resin. They may appear as very light particles in thick sections observed by light microscopy. They lie in large free clusters in the cytoplasm, away from other organelles, and have no associated membrane.
See Gut granule
|
| Golgi |
Golgi apparatus (S)
Golgi body (S)
Dictyosome (S) |
|
Large membranous organelles lying in the cell cytoplasm, often clustered near the nucleus and the RER; more numerous in cells with high level of secretory activity. |
| Gonad |
|
|
A specialized tissue of the reproductive tract that produces the male or female gametes. In hermaphroditic nematodes, one gonad, or “ovotestis”, can produce both male gametes and female gametes in succession. The reproductive tract in the hermaphrodite has two equivalent gonad “arms”, while the male has a one-armed gonad. Each gonad arm bends in the middle (a reflexion, often called the "loop"). As it grows longer, that portion lying closer to the gonopore (vulva or cloaca) is termed the “proximal arm”, while the “distal arm” lies further away with the extreme end named the "distal tip”. Each mature gonad arm generally consists of a core of germline cells surrounded by an outer somatic epithelium (somatic gonad), comprised of the gonad sheath (ovary), spermatheca, uterus (oviduct) and vulva in the hermaphrodite, or gonadal sheath, seminal vesicle and cloaca in the male.
See Distal tip cell
See Ovary
See Oviduct
See Sheath
See Seminal vesicle
See Testis
See Uterus
See Vas deferens
|
| Gonadal leader |
Distal tip cell (S)
Linker cell (S)
|
|
The somatic gonad cell, also known as the distal tip cell (in the hermaphrodite) or the linker cell (in the male), which guides the outgrowth of the gonad primordium into an extended tube-shaped structure. It crawls along the bodywall, pulling or leading the developing gonadal cells which are behind it, while also secreting a signal which causes nearby germ cells to remain in mitosis. The two-armed gonad in the hermaphrodite has two leader cells, whereas the one-armed male gonad has one leader cell.
See Distal tip cell
See Linker cell |
| Gonadal migration |
|
|
This term refers to the gross movement of the whole structure of the gonad along the bodywall, behind the gonadal leader cell and away from the developing vulva. The gonad primordium begins as a cluster of four blast cells on the ventral side of the midbody in the embryo. During larval stages these cells begin to divide and create a much larger mass of cells which then must be shaped into a long tubular epithelium surrounding an extended mass of germ cells. Eventually, each gonad arm migrates away from the vulva, turns dorsally and then reflects back towards the midpoint along the bodywall. See animation.
See Distal tip cell
See Reflexion
|
| Gonadal primordium |
|
|
A small cluster of cells in the embryo and young larva that begin to develop into the gonad during the late L1 stage. Initially the primordium consists of just four cells: Z1and Z4 give rise to the somatic cells of gonad, while Z2 and Z3 give rise to the germline itself. Their subsequent lineages are sex-specific and occur during larval development (Kimble and Hirsh, 1979). |
| Gonadogenesis |
|
|
The developmental process through which the adult gonad is formed from an initial set of four blast cells, Z1, Z2, Z3, Z4, born in the early embryo (Kimble and Hirsh, 1979). |
| Gonad sheath |
Cellular sheath (S)
Somatic sheath (S) |
|
A set of five cell pairs which form a thin continuous tube surrounding the proximal portions of the ovary and covering the maturing oocytes from the spermatheca to the loop region and over the near portion of the distal gonad arm. These cells are tightly joined to each other and to the spermatheca by adherens junctions, have fenestrations which permit yolk to pass from the pseudocoelom to maturing oocytes, and communicate directly to mature oocytes by gap junctions. The most distal sheath cell pair extends long filapodia over the distal germline to stretch the sheath over the distal arm (Hall et al., 1999).
See Contractile gonad sheath of the hermaphrodite |
| Gonochoristic |
Gonochoric (S) |
|
The method of reproduction in which male and female germ cells are provided by two sexes and fertilized within the female. As opposed to hermaphroditism or parthogenesis.
See Dioecious
See Hermaphrodite |
| Gonoduct |
|
|
An archaic term referring collectively to those passageways which carry germ cells or fertilized oocytes within the body, e.g. the gonad sheath, uterus, and vulva in the hermaphrodite, and the vas deferens in the male.
See Reproductive tract |
| Gonopore |
|
|
A narrow body opening through which germ cells and/or embryos are released into the exterior.
See Cloaca
See Vulva |
| GPI-linkage |
GPI-anchor
GPI-anchorage
|
|
A protein that becomes attached to the plasma membrane via a specific covalent linker involving glycosylphosphotidylinositol (GPI) rather than via a transmembrane protein sequence. |
| Granule |
|
|
A dark-staining object in the cell cytoplasm or in the nucleoplasm. These usually correspond to a membrane-bound, secretory or storage vesicle (a yolk granule, for instance), but could also refer to a compact dense structure without a membranous coat (such as a P granule).
Alternately, an “internal granule” can be a small dark object lying within a larger clear vesicle or vacuole (Leung et al., 1999).
See Glycogen
See P granules
See Secretory granule |
| Gravid |
|
|
Generally means carrying eggs or unborn young (being pregnant). In C. elegans refers to an adult hermaphrodite full of fertilized eggs and embryos.
See Post-reproductive
|
| Grinder |
Teeth (S)
Cuticle knobs (S)
Esophageal valve (S)
|
|
A highly specialized cuticle region of the pharynx inside the lumen of the second bulb, secreted by the lumenal surface of the pm6 and pm7 muscle cells (Avery and Thomas, 1997). The grinder appears to physically break open bacteria to aid in their digestion. These irregular teeth are probably scraped against one another by motor actions of radial muscles (pm6) and longitudinal muscles (pm7).
Some authors interpret this structure as a valve rather than a feeding apparatus (Nicholas, 1975). In this interpretation, the interlocking teeth are essential to regulate pressure changes within the lumen and to seal the contents from backflow out the buccal opening during pumping. Nicholas (1975) suggests that high turgor pressure will normally act to shut down the lumen, but that coordinated opening of first the anterior and then the posterior haustrulum (by muscle contractions in the second bulb) permits one-way passage of food particles via a click-shut mechanism of the valve’s teeth.
See Haustrulum
|
| Ground substance |
Ground material (S)
Matrix (S) |
|
Material which evenly fills the cytoplasmic space (cytosol) or extracellular space in a proscribed region. It may be of medium density or very electron dense. It often forms a distinct background for other darker or lighter objects (with more distinctive borders or membranes) within the same space. |
| Growth cone |
|
|
An enlargement of the distal end of a growing axon, from which several lamellipodia and thin filapodia extend along the bodywall to lead the growth cone in a particular direction, based upon the reception of guidance cues from the local environment. Receptors expressed in the growth cone membrane are thought to transduce attractive and repulsive cues which either promote further filapodial (or lamellipodial) extensions towards the signal source, or cause filapodial collapse and turning away. The shape of the growth cone is very plastic, based upon actin-based cytoskeletal dynamics, whereas the trailing axon shape is more stable, based mostly upon microtubule extension to shape its structure.
See Guidance cue
See Filapodium
See Follower axon
See Pioneer axon
|
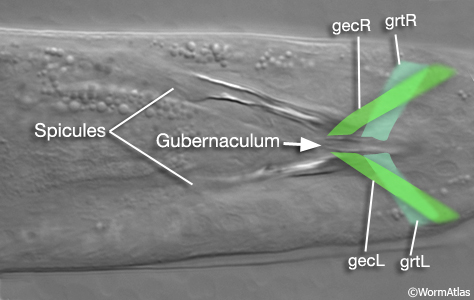
| Gubernaculum |
Spicular pouch (S) |
|
A sclerotized cuticular structure at the base of the male tail fan which houses the retracted spicules and guides the eversion of the spicules through the anus. It also features a small sensory “hook” at the base of the spicules which may be involved in triggering their eversion. In some references, this structure also includes the cloacal opening, and the ventral side of the tail fan.
Alternately, Sulston et al. (1980) uses this term to refer only to the sclerotized cuticle itself; but this usage seems non-standard. |
| Gubernacular muscles |
|
|
A specialized set of sex-specific muscles attached to the gubernaculum in the male tail which are required to change the shape of the gubernaculum during male mating behavior. These two bilateral muscle pairs are the gubernacular erectors (gecL/R) and retractors (grtL/R). |
| Guidance cue |
|
|
Items along the trajectory of a moving object (whole animal, cell, tissue, organelle, or cell part) that are used in determining changes in direction. They can consist of physical barriers to continued progress, differences in adhesive qualities of the substrate, or molecular signals provided into the medium as soluble factors or membrane-bound moieties. Molecular cues might be arrayed as graded signals along the trajectory, or in broad stripes, and may be attractive or repulsive to the moving object. |
| Guidepost cell |
|
|
This term is used in reference to a common mechanism of axon guidance. The guidepost cell is thought to present a “guidance cue” on its plasma membrane or secrete the cue into its surround (perhaps onto the local basal lamina). The cue molecules act to attract or repel the advance of certain axon growth cones which express a receptor for that guidance cue. |
| Gustatory plasticity |
|
|
A form of chemotaxis behavior in which pre-exposure to high salt changes the animal’s preference to intermediate salt concentrations (Jansen et al., 2002). |
| Gut |
Intestine (S)
|
|
A general term which may refer both to the intestine and to related muscles and valves which regulate intake or outflow of its contents. |
Gut granule
|
|
|
A nonspecific term which may encompass a variety of different inclusions within the intestinal cytoplasm, including birefringent crystalline objects and other dark-staining endosomes. Their contents might include glycogen granules (polysaccharide storage), rhabditin (carbohydrate storage), yolk granules (lipid and protein), and concentrated waste products.
In some naming schemes, the more highly birefringent crystals may be termed as “spheroids” or “spherocrystals” while dense-staining, less birefringent objects are categorized as gut granules, and non-staining polysaccharide deposits are categorized separately as glycogen granules.
See Glycogen
See Rhabditin
See Yolk
|
| Gymnostom |
Prostom (S)
|
|
The middle portion of the buccal capsule, bordered by the arcade cells. This region lies just anterior to the stegostom and posterior to the cheilostom (Dolinski et al., 1998). |


{kind=link}