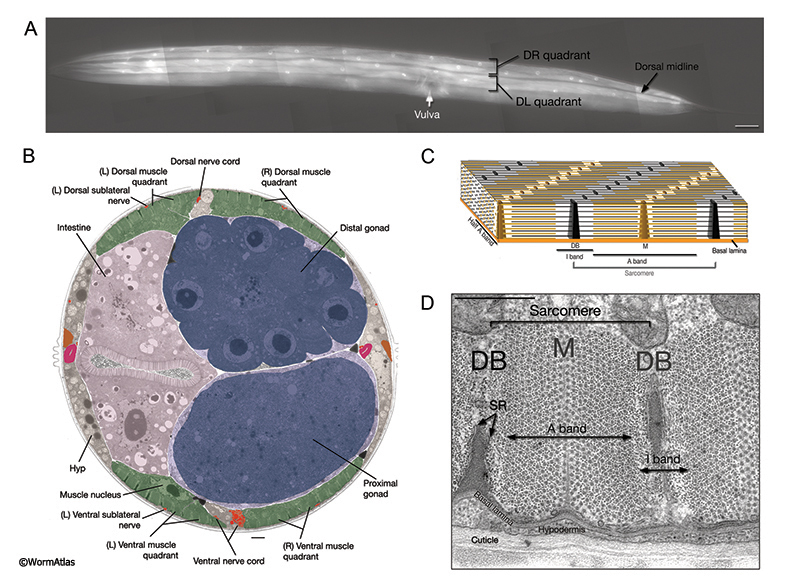
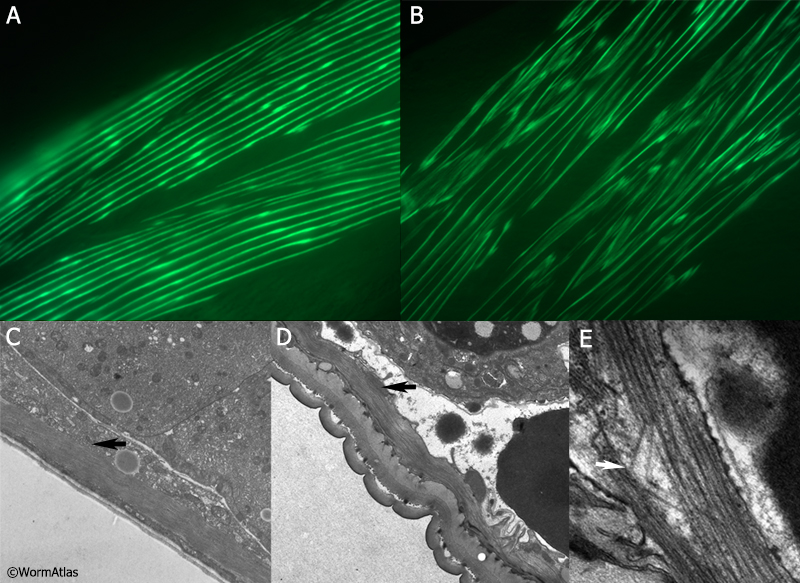
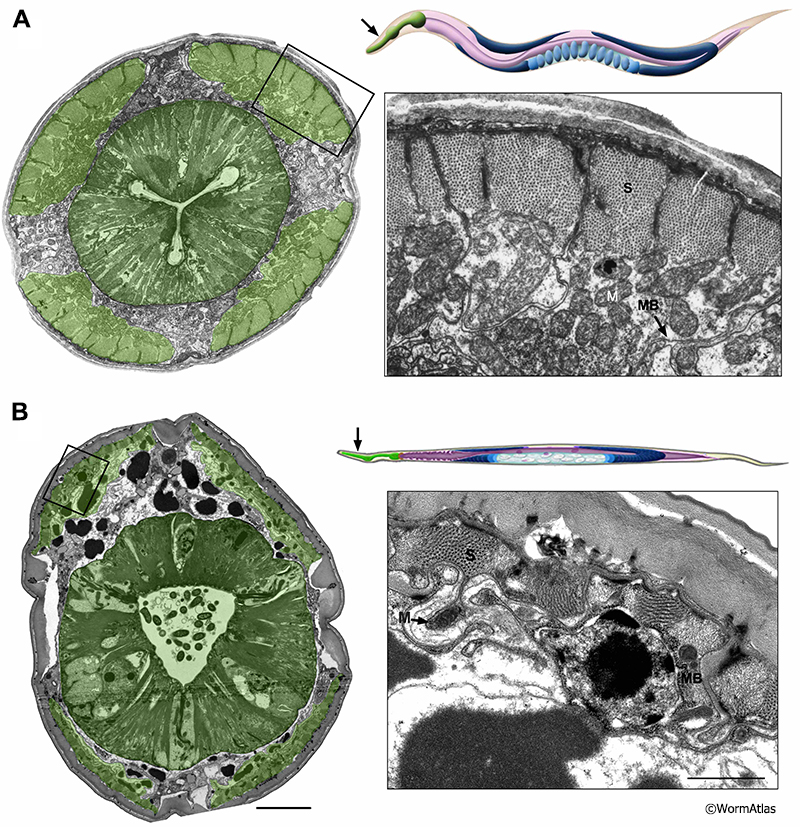
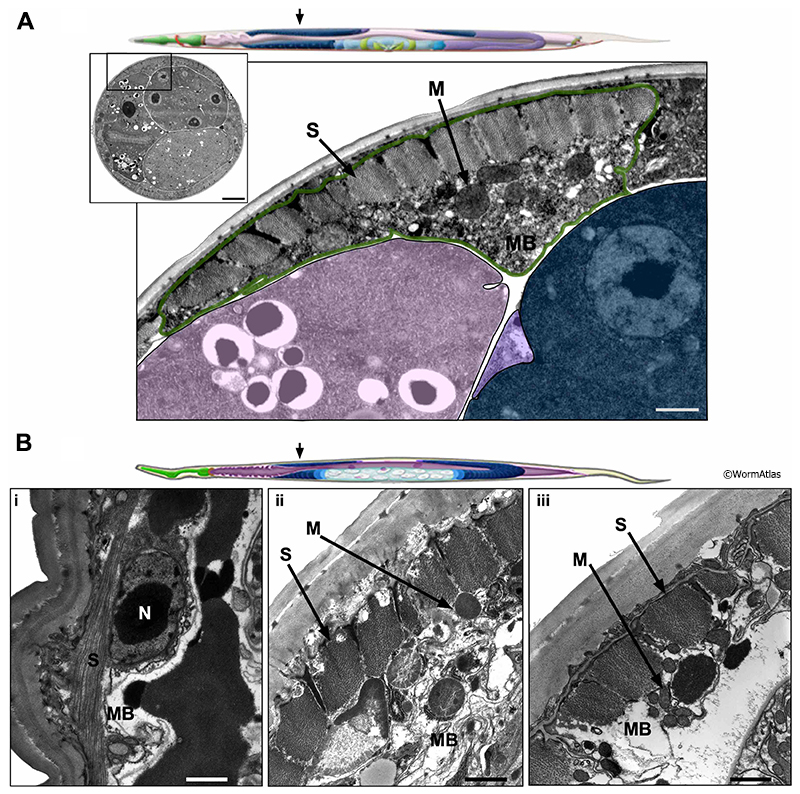
4 References
Alway, S.E., Mohamed, J.S. and Myers, M.J. 2017. Mitochondria initiate and regulate sarcopenia. Exerc. Sport. Sci. Rev. 45: 58-69. doi: 10.1249/JES.0000000000000101
Bansal, A., Zhu, L.J., Yen, K. and Tissenbaum, H.A. 2015. Uncoupling lifespan and healthspan in Caenorhabditis elegans longevity mutants. PNAS. 112: E277-E286. doi: 10.1073/pnas.1412192112
Bolanowski, M.A., Russell, R.L. and Jacobson, L.A. 1981. Quantitative measures of aging in the nematode Caenorhabditis elegans. I. Population and longitudinal studies of two behavioral parameters. Mech. Ageing Dev. 15: 279-95. doi: 10.1016/0047-6374(81)90136-6 Article-pdf
Chang, J.T. and Hansen, M. 2018. Age-associated and tissue-specific decline in autophagic activity in the nematode C. elegans. Autophagy 14: 1276-1277. doi: 10.1080/15548627.2018.1445914
Chuang, H.S., Kuo, W.J., Lee, C.L., Chu, I.H. and Chen, C.S. 2016. Exercise in an electrotactic flow chamber ameliorates age-related degeneration in Caenorhabditis elegans. Sci. Rep. 6: 28064. doi: 10.1038/srep28064
Coen, P.M., Musci, R.V., Hinkley, J.M. and Miller, B.F. 2019. Mitochondria as a target for mitigating sarcopenia. Front. Physiol. 9: 1883. doi: 10.3389/fphys.2018.01883
Croll, N.A., Smith, J.M. and Zuckerman, B.M. 1977. The aging process of the nematode Caenorhabditis elegans in bacterial and axenic culture. Exp. Aging Res. 3: 175–189. doi: 10.1080/03610737708257101 Article-pdf
Dhondt, I., Verschuuren, C., Zecic, A., Loier, T., Braeckman, B.P. and De Vos, W.H. 2021. Prediction of biological age by morphological staining of sarcopenia in Caenorhabditis elegans. Dis. Model Mech. 14: dmm049169. doi: 10.1242/dmm.049169
Duhon, S.A. and Johnson, T.E. 1995. Movement as an index of vitality: comparing wild type and the age-1 mutant of Caenorhabditis elegans. J. Gerontol. Biol. Sci. 50: B254-B261. doi: 10.1093/gerona/50a.5.b254
Gaffney, C.J., Pollard, A., Barratt, T.F., Constantin-Teodosiu, D., Greenhaff, P.L. and Szewczyk, N.J. 2018. Greater loss of mitochondrial function with ageing is associated with earlier onset of sarcopenia in C. elegans. Aging 10: 3382-3396. doi: 10.18632/aging.101654
Garcia-Casas, P., Alvarez-Illera, P., Gomez-Orte, E., Cabello, J., Fonteriz, R.I., Montero, M. and Alvarez, J. 2021. The mitochondrial Na+/Ca2+ exchanger inhibitor CGP37157 preserves muscle structure and function to increase lifespan and healthspan in Caenorhabditis elegans. Front. Pharmacol. 12: 695687. doi: 10.3389/fphar.2021.695687
Garigan, D., Hsu, A.L., Fraser, A.G., Kamath, R.S., Ahringer, J. and Kenyon, C. 2002. Genetic analysis of tissue aging in Caenorhabditis elegans: a role for heat-shock factor and bacterial proliferation. Genetics 161:1101–1112. doi: 10.1093/genetics/161.3.1101 Article-pdf
Glenn, C.F., Chow, D.K., David, L., Cooke, C.A., Gami, M.S., Iser, W.B., Hanselman, K.B., Goldberg, I.G. and Wolkow, C.A. 2004. Behavioral deficits during early stages of aging in Caenorhabditis elegans result from locomotory deficits possibly linked to muscle frailty. J. Gerontol. 59A: 1251-60. doi: 10.1093/gerona/59.12.1251 Article-pdf
Haithcock, E., Dayani, Y., Neufeld, E., Zahand, A.J., Feinstein, N., Mattout, A., Gruenbaum, Y. and Liu, J. 2005. Age-related changes of nuclear architecture in Caenorhabditis elegans. PNAS 102: 16690-16695. doi: 10.1073/pnas.0506955102
Hartman, J.H., Smith, L.L., Gordon, K.L., Laranjeiro, R., Driscoll, M., Sherwood, D.R. and Meyer, J.N. 2018. Swimming exercise and transient food deprivation in Caenorhabditis elegans promote mitochondrial maintenance and protect against chemical-induced mitotoxicity. Sci. Rep. 8: 8359. doi: 10.1038/s41598-018-26552-9
Herndon, L.A., Schmeissner, P.J., Dudaronek, J.M., Brown, P.A., Listner, K.M., Sakano, Y., Paupard, M.C., Hall, D.H. and Driscoll, M. 2002. Stochastic and genetic factors influence tissue-specific decline in ageing C. elegans. Nature 419: 808-814. doi: 10.1038/nature01135 Article-pdf
Herndon, L.A., Wolkow, C.A., Driscoll, M. and Hall, D.H. 2017. Effects of ageing on the basic biology and anatomy of C. elegans. In Ageing: lessions from C. elegans. (ed Olsen, A. and Gill, M.). Chapter 2. pp. 9-39. Springer International, Switzerland. doi: 10.1007/978-3-319-44703-2_2 Article-pdf
Huang, C., Xiong, C. and Kornfeld, K. 2004. Measurements of age-related changes of physiological processes that predict lifespan of Caenorhabditis elegans. Proc. Natl. Acad. Sci. U S A. 101:8084–8089. doi: 10.1073/pnas.0400848101 Article-pdf
Ibáñez-Ventoso, C., Herrera, C., Chen, E., Motto, D. and Driscoll, M. 2016. Automated analysis of C. elegans swim behavior using CeleST software. J. Vis. Exp. 118: 54359. doi: 10.3791/54359
Johnson, T.E. 1987. Aging can be genetically dissected into component processes using long-lived lines of Caenorhabditis elegans. Proc. Natl. Acad. Sci. USA 84: 3777-81. doi: 10.1073/pnas.84.11.3777
Johnston, J., Iser, W.B., Chow, D.K, Goldberg, I.G. and Wolkow, C.A. 2008. Quantitative image analysis reveals distinct structural transitions during aging in Caenorhabditis elegans tissues. PLoS ONE. 3: e2821. doi: 10.1371/journal.pone.0002821 Article-pdf
Kern, A., Ackermann, B., Clement, A.M., Duerk, H. and Behl, C. 2010. HSF-1-controlled and age-associated chaperone capacity in neurons and muscle cells of C. elegans. PLoS ONE. 5: e8568. doi: 10.1371/journal.pone.0008568
Koyuncu, S., Loureiro, R., Lee, H.J., Wagle, P., Krueger, M. and Vilchez, D. 2021. Rewiring of the ubiquitinated proteome determines ageing in C. elegans. Nature. 596: 285-290. doi: 10.1038/s41586-021-03781-z
Laranjeiro, R., Harinath, G., Burke, D., Braeckman, B.P. and Driscoll, M. 2017. Single swim sessions in C. elegans induce key features of mammalian exercise. BMC Biol. 15: 30. doi: 10.1186/s12915-017-0368-4
Laranjeiro, R., Harinath, G., Hewitt, J.E., Hartman, J.H., Royal, M.A., Meyer, J.N., Vanapalli, S.A. and Driscoll, M. 2019. Swim exercise in Caenorhabditis elegans extends neuromuscular and gut healthspan, enhances learning ability, and protects against neurodegeneration. Proc. Natl. Acad. Sci. USA. 116: 23829-23839. doi: 10.1073/pnas.1909210116
Liu, J., Zhang, B., Lei, H., Feng, Z., Liu, J., Hsu, A.L and Xu, X.Z. 2013. Functional aging in the nervous system contributes to age-dependent motor activity decline in C. elegans. Cell Metab. 18: 392-402. doi: 10.1016/j.cmet.2013.08.007
Mallick, A., Ranawade, A., van den Berg, W. and Gupta, B.P. 2020. Axin-mediated regulation of lifespan and muscle health in C. elegans requires AMPK-FOXO signaling. iScience. 23: 101843. doi: 10.1016/j.isci.2020.101843
Mergoud dit Lamarche, A., Molin, L., Pierson, L., Mariol, M.-C., Bessereau, J.-L., Gieseler, K. and Solari, F. 2018. UNC-120/SRF independently controls muscle aging and lifespan in Caenorhabditis elegans. Aging Cell 17: 12713. doi: 10.1111/acel.12713
Regmi, S.G., Rolland, S.G. and Conradt, B. 2014. Age-dependent changes in mitochondrial morphology and volume are not predictors of lifespan. Aging. 6: 118-130. doi: 10.18632/aging.100639
Restif, C., Ibáñez-Ventoso, C., Vora, M.M., Guo, S., Metaxas, D. and Driscoll, M. 2014. CeleST: computer vision software for quantitative analysis of C. elegans swim behavior reveals novel features of locomotion. PLoS Comput. Biol. 10: e1003702. doi: 10.1371/journal.pcbi.1003702
Sudevan, S., Takiura, M., Kubota, Y., Higashitani, N., Cooke, M., Ellwood, R.A., Etheridge, T., Szewczyk, N.J. and Higashitani, A. 2019. Mitochondrial dysfunction causes Ca2+ overload and ECM degradation-mediated muscle damage in C. elegans. FASEB J. 33: 9540-9550. doi: 10.1096/fj.201802298R
Tiku, V., Jain, C., Raz, Y., Nakamura, S., Heestand, B., Liu, W., Spth, M., Suchiman, H.E.D., Mller, R.U., Slagboom, P.E., Partridge, L. and Antebi, A. 2017. Small nucleoli are a cellular hallmark of longevity. Nat Commun. 8: 16083. doi: 10.1038/ncomms16083
Wang, H., Webster, P., Chen, L. and Fisher, A.L. 2019. Cell-autonomous and non-autonomous roles of daf-16 in muscle function and mitochondrial capacity in aging C. elegans. Aging. 11: 2295-2311. doi: 10.18632/aging.101914
Weir, H.J., Yao, P., Huynh. F.K., Escoubas, C.C., Goncalves, R.L., Burkewitz, K., Laboy, R., Hirschey, M.D. and Mair, W.B. 2017. Dietary restriction and AMPK increase lifespan via mitochondrial network and peroxisome remodeling. Cell Metab. 26: 884-96. doi: 10.1016/j.cmet.2017.09.024
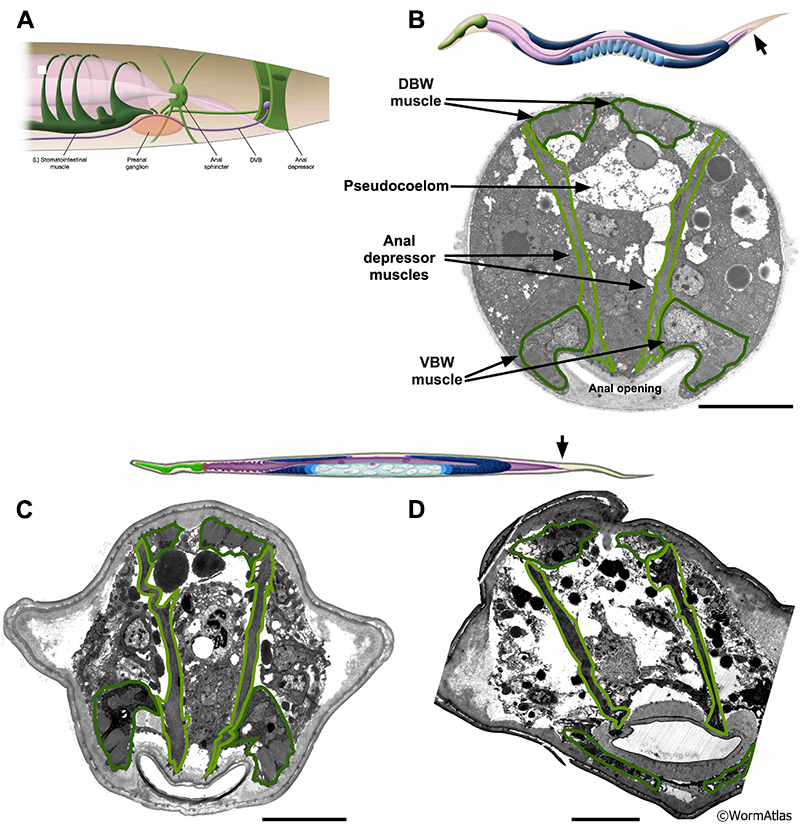
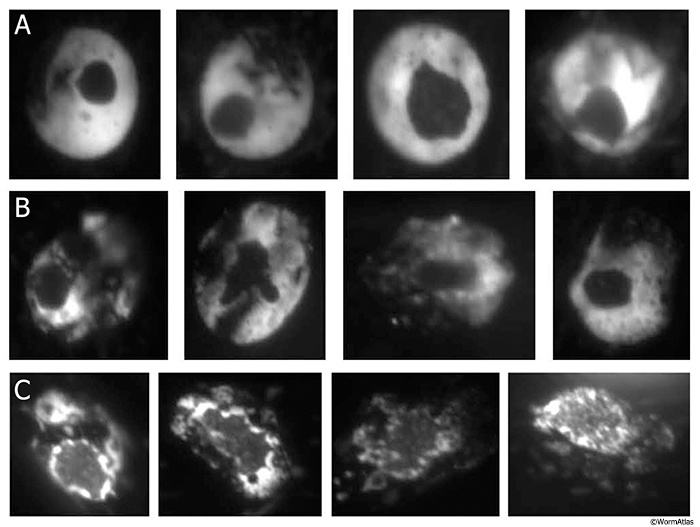
* Description of Behavioral Classes (A, B, C) as described in Herndon et al., 2002
To characterize aging phenotypes, age-synchronized individual worms were scored both for spontaneous movement and for response to prodding with a wire over the course of their lifespan. Three distinct classes representing behavioral phenotypes were established. Animals that move constantly and make sinusoidal tracks were designated as class A. Class B animals mainly move when prodded. When they move it is with uncoordinated motion, leaving non-sinusoidal tracks. Class C animals do not move forward or backward, even upon prodding, but do show head and/or tail movement and twitch in response to touch. All animals begin adulthood in class A. Class B animals appear around days 6-7 of adulthood and class C around day 9-10 (at 20oC). At later ages, animals representing all classes can be found within the same population and it was found that the behavioral class type was the better predictor of life expectancy than chronological age (Herndon et al., 2002). Due to the stochastic nature of the aging process in an individual nematode, these classifications only reflect ongoing changes in nerve and muscle, while other tissues can show very different age-related effects within one behavioral class, declining faster or remaining healthy much longer.
|

 Click pictures for new window with figure and legend, click again for high resolution image
Click pictures for new window with figure and legend, click again for high resolution image