| PART/CELL
NAME |
ABBREVIATION
SYNONYMS (S)
ANTONYMS (A) |
LINEAGE |
DESCRIPTION |
C |
C blastomere (S)
C founder cell (S) |
P0.ppa |
An embryonic founder cell which
is born dorsally and whose daughter cells give rise to dorsal hypodermis,
some somatic muscle cells and several neurons.
In earlier publications,"C"
may also refer to the postembryonic blast C cell (C ectoblast) which was
later renamed "Y". |
| C blastomere |
|
|
C founder
cell |
| "c"
neuron |
AWC cell (S) |
|
Amphid wing "c" cells, neurons having ciliated sheet-like sensory
endings closely associated with amphid sheath
See AW
(Amphid wing) cell
|
| "C"
neuron |
CEP cell (S) |
|
See CEP
cell |
| CA
cell |
CA1
CA2
CA3
CA4
CA5
CA6
CA7
CA8
CA9 |
P3.aapa
P4.aapa
P5.aapa
P6.aapa
P7.aapa
P8.aapa
P9.aapa
P10.aapa
P11.aapa |
Male specific cell
Male specific cell
Male specific cell
Male specific neuron, innervates dorsal muscles
Male specific neuron, innervates dorsal muscles
Male specific neuron, innervates dorsal muscles
Male specific neuron, innervates dorsal muscles
Male specific cells in VC, neuron-like, but lack synapses
Male specific cells in VC, neuron-like, but lack synapses
|
| Caenorhabditis
brenneri |
|
|
Previously unnamed strain PB2801 was renamed in honor of Syndey Brenner. While closely related to C. elegans and C. briggsae, and having similar biological characteristics, adults are either male or female, not hermaphrodites. Sequencing of the genome is ongoing.
See Phylogeny
|
| Caenorhabditis
briggsae |
|
|
A commonly
studied species of nematode whose anatomical features are extremely similar
to Caenorhabditis elegans, and whose genetic sequences are being
compared in exacting detail against the genome of C. elegans. C.
briggsae and C. elegans cannot interbreed to create fertile progeny,
but are otherwise extremely similar in behavior and ecology; they are likely
to be very close relatives evolutionarily (Nigon
and Dougherty, 1949; Baird
et al., 1992).
See Phylogeny
|
| Caenorhabditis
elegans |
|
|
A species
of nematode, or roundworm, that has been adopted as a model organism for
the study of genetics, cell biology, development, and neuroscience. Two
different wild strains, Bristol and Bergerac, were initially adopted by Nigon
and Dougherty (1949) and by Brenner
(1974) as the basis for scientific studies, although additional wild-caught
strains have since been collected around the world for comparisons of genetic
diversity. The sequence of the entire genome was published in 1998 (with the last gaps finished in 2002) making C. elegans the first multi-cellular eukarote in which every base is known (C. elegans Sequencing Consortium, 1998). |
| Caenorhabditis
japonica |
|
|
A species
of rhabditid nematode closely related evolutionarily to Caenorhabditis
elegans and Caenorhabditis briggsae. Like C. remanei, C. japonica does not
form hermaphrodites, but utilizes a male/female sexual mating system. Additionally, it appears that unlike C. elegans, C. japonica may enter the dauer stage regardless of food and environmental conditions. This species is usually found in close association with stink bugs. Full sequencing of the genome is currently in the initial stages. |
| Caenorhabditis
remanei |
|
|
A species
of rhabditid nematode closely related evolutionarily to Caenorhabditis
elegans and Caenorhabditis briggsae. C. remanei does not
form hermaphrodites, but utilizes a male/female sexual mating system. For
research purposes, most widely used strains of C. remanei are EM464 and SB146. Note that the strain CB5161, which was called C. remanei for some time, is actually a different species (and now believed to be C. brenneri). Full sequencing of the genome is currently in the initial stages. |
| CAN cell |
CANL
CANR |
ABalapaaapa
ABalappappa |
A pair
of neurons with processes that run along excretory canals and within canal
associated nerves. Make no synapses, but essential for survival. |
| Canal
associated nerve |
|
|
A pair
of longitudinal nerve tracts that run alongside the excretory canals on
the right and left side of the animal. Processes from CAN, BDU, PVD and ALA form this nerve. |
| Canaliculus/Canaliculi (/plu) |
|
|
A series
of thin membranous collecting channels, closed at their distal ends, which
feed into lumen of each excretory canal. The canaliculi can differ in shape
depending on the status of the animal, perhaps reflecting changes in osmolarity
or secretory activity. Thus the canaliculi can be 1) smooth, narrow and
sinuous, or 2) undulating in diameter to form a series of beads on a chain,
or 3) disconnected to form a series of larger spherical vesicles. |
| Canonical
allele |
|
|
The original
isolate of a mutation whose properties define the basis for recognizing
this strain as a separate class of mutant. (e.g. let-7(mn122) and unc-35(e259)). The severity of the phenotype of this allele is used
as a measure against which the phenotypes of later alleles are compared. |
| Cap |
Cap cell (S)
Socket cell (S) |
|
1) Older name for socket cell.
2) Can also refer to a large
piece of shed cuticle that had originally covered the head. From the cap’s
dimensions, it has been suggested that secretions from the excretory system
might provide a molting enzyme that works to weaken this piece of cuticle
near the excretory pore at the back of the head in some species (Chitwood
and Chitwood, 1951; Bird and Bird, 1991). In C. elegans, laser
ablation experiments to remove the excretory system have not inhibited
the molting process (Singh
and Sulston, 1978).
See Socket cell
|
| Cardia |
|
|
See Pharyngeal-intestinal
valve |
Cauda/Caudal |
|
|
Tail/of a tail, relating to
a tail. Located at or directed towards the posterior part of the body. |
| Caudal
alae |
|
|
The lateral specialization
of the cuticle in the male tail, more commonly known as the fan.
See Fan
|
| Caudal glands |
|
|
These are found in "Adenophorea" only.
There are 3 caudal glands with a common or separate openings at the tip
of the tail. The secretion is used to attach the worm to the substrate. |
| Caudal
papillae |
Male
tail rays (S) |
|
See Ray |
| Caudalids (archaic) |
Lumbar comissures (S) |
|
The nerve connections between the lumbar ganglia and the pre-anal ganglion, which must circumvent the ventral opening of the anus. |
| Caveola
/ Caveolae (/plu) |
|
|
Vesicular organelles which
are a specialized feature of some plasma membranes. They could be the
basis of endocytosis or other forms of membrane-trafficking in some nematode
cell types. Caveolae are notably smaller and less electron dense than
coated vesicles 50-100 nm for caveolae, 200-300 nm diameter for coated
vesicles, according to Razani
and Lisanti (2001). Caveolin proteins are principal components of
these organelles.
A caveolin-1 mutant in C. elegans blocks cell cycle progression
(Scheel
et al., 1999).
See Coated vesicle
|
ccDL herm
ccDR herm |
|
M.dlpa
M.drpa |
A pair
of postembryonic coelomocytes in hermaphrodite. |
| ccD male |
|
M.dlpappp |
Single
postembryonic coelomocyte in male. |
ccAL
ccAR
ccPL
ccPR |
|
MSapapaaa
MSppapaaa
MSapapaap
MSppapaap |
Embryonic
coelomocytes |
| Cecum/Ceca
(/plu) |
|
|
An outpocketing
(diverticulum) or blind ending of the lumen of the intestine, usually forming
a side branch of the lumen proper. In C. elegans, the anterior intestinal
lumen between the first four cells is quite large in volume, but no distinctive
side branches form there.
Ceca are common in some other nematode species,
especially at the anterior limit, and may form along the length of the intestine
of some C. elegans mutants. |
| Ced |
|
|
CEll Death abnormality
phenotype or mutation.
See Cell death
|
| Cell
body |
Soma
(S) |
|
The largest
portion of a cell, generally surrounding the nucleus and containing abundant
cytoplasm and organelles. |
| Cell
death |
|
|
There are a number of distinct
mechanisms by which individual cells within the organism can die, generally
without direct death of the whole animal. In C. elegans, many cells
die via apoptosis, or programmed cell death, soon after their birth (Conradt and Xue, 2005). In
a variety of disease mechanisms, in old age, in some mutant phenotypes,
and after toxic treatments, various cells may be induced to die by alternate
mechanisms, which include necrosis (Syntichaki and Tavernarakis, 2002). These pathways are distinct, but share some components and cellular machinery. The morphological
features of these various modes of death are in some ways distinctive,
but dying cells can often show mixed characters. The corpses of dead cells
rarely linger, but are quickly engulfed and phagocytosed by neighboring
cells.
See Apoptosis
See Necrosis
See Autophagy
See Programmed cell death
See Engulfment
|
| Cell
fusion |
|
|
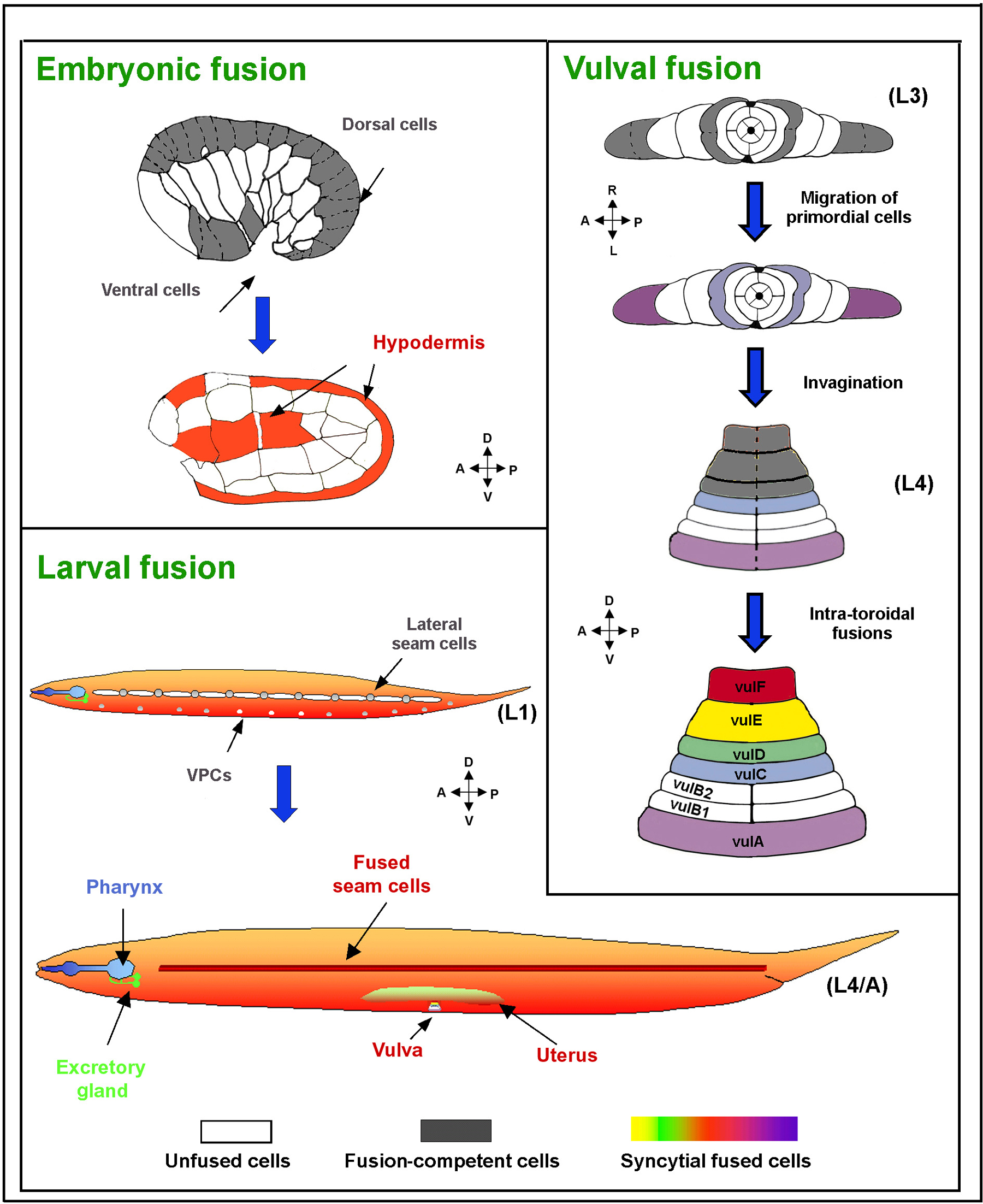
A common feature in C. elegans tissues, in which the lateral borders of two or more cells become tightly
fused and then open to form a patent channel between the cells (Podbilewicz
and White, 1994; Podbilelwicz, 2006). In some cases, the point of fusion remains quite
small, and fragments of the former cell border can be retained, such that
the cells seem virtually separate (as in pharyngeal muscle cell pairs).
In other cases, all vestiges of the former border are removed, cell nuclei
migrate across from one zone to the next, and the combined syncytial structure
shows little resemblance to the original chain of cells (as in the hyp7
syncytium). In the germline, all young germ cells are fused to a common
rachis and may share intracellular components with the rachis, yet these
cells mature independently and eventually separate from the rachis (reversing
In C. elegans, syncytial
cells include some of the hypodermal cells, excretory gland cell, vulva
epithelium, uterus epithelium, pharyngeal muscles, pharyngeal epithelial
marginal cell, pharyngeal g1 gland cell, ant and post arcade, tail spike
cells and seam cells.
See Rachis
See Syncytium
|
| Cell
intercalation |
|
|
A developmental
process that results in the rearrangement of several C. elegans tissues
in which cells migrate for short distances from their place of birth to
surround a developing epithelial structure, as in the intestine and vulva,
or to form a discrete chain of cells such as the ventral nerve cord and embryonic hypodermis.
Intercalation can be used to describe the positions of excess cells (or
structures) that come to lie within a row of other cells due to changes
in cell fate due to mutation or developmental anomalies. |
| Cell
invasion |
|
|
See Invasion |
| Cell
lineage |
|
|
The pattern
of cell divisions and cell fates produced by an ancestral blast cell. This
series of cell divisions is highly stereotyped in C. elegans (See Lineage tree),
so that most dividing cells have reproducible axes of their division planes
(e.g. dorsal/ventral, anterior/posterior, etc), and reproducible timing
in the divisions and asymmetries in the relative size of the two daughter
cells. Segregation of nuclear determinants within the two daughters is also
reproducible resulting in consistent cell fates among the daughters from
animal to animal. Cell lineages are often depicted graphically as decision
trees. |
| Cell
migration |
|
|
Some
cells are born at some distance from their terminal locations as mature
cells, and must crawl in amoeboid fashion within the body cavity to reach
their eventual destination. They follow molecular signals expressed by cells
along the route of their migration or by cells within their target tissue.
During gastrulation, many young cells shift positions within the expanding
embryo to form an internal layer of cells. The distal tip cells differentiate
and then begin to crawl within the body cavity to guide outgrowth of the
gonad. The hypodermal cells of the male tail migrate to lead the retraction
(migration) of other cell types from the extreme tail tip prior to morphogenesis
of the posterior fan and rays.
One differentiated cell type, the gonad sheath
cell (1st pair only) in the mature adult gonad appears to crawl continuously
by filopodial extension to try to cover the exposed surface of the dividing
germline. Some sensory neurons which are born at the surface of the body,
later migrate inward and towards the central ganglia, leaving behind a sensory
dendrite attached to the bodywall. Many larval cells make shorter migrations
in tissues as they reposition themselves to form epithelial layers, including
the developing intestine. |
| Cellular sheath |
|
|
See Gonad
sheath |
| Cellularization |
|
|
Process by which maturing germ
cells, which are initially connected via the rachis within the distal
gonad arm, break free from the rachis to continue the final stages of
development, either as individual spermatocytes or oocytes, or to undergo
programmed cell death.
See Cell fusion
See Nurse cells
See Rachis
|
| CEM cell |
CEMDL
CEMDR
CEMVL
CEMVR
Cephalic companion cell (S) |
ABplaaaaaap
ABarpapaaap
ABplpaapapp
ABprpaapapp |
A sensory neuron cell type
found in the head of males but not hermaphrodites (these cells die in the hermaphrodite
embryo). Each CEM shares the same sheath cell as the CEP neuron in the
four dorsal and ventral lips, and forms a sense ending which opens separately
to the exterior through the cuticle (See Cephalic Sensilla). They may mediate male chemotaxis towards hermaphrodites. |
| Central
cells |
|
|
See Wishbone cells |
Central
cylinder |
|
|
See Apical ring |
| Central element |
|
|
A specific portion of the chromosomal pair during synapsis that forms part of the synaptonemal complex and runs between the lateral elements. Ladder-like in structure, it has two longitudinal components and regularly spaced transverse components that bridge the longitudinal components and may be subdivided into 3-4 layers (Schmekel and Daneholt, 1995).
See Axial element
See Lateral element
See Synaptonemal complex |
Centration |
|
|
A coordinated movement during prophase of the two centrosomes and the two pronuclei towards the center of the one cell embryo (Gönczy and Rose, 2005) |
| Centriole |
|
|
A small cylindrical structure, usually found in pairs, that are found at each pole of the mitotic spindle in animal cells and some other eucaryotes. Centrioles lie perpendicular to one another to form the centrosome and organize the mitotic spindle for cell division. While in most animals, centrioles are composed of nine triplet microtubules, in C. elegans the centriole has a nine singlet conformation (for more detail see Oegema and Hyman, 2006).
Centrioles are also found distally in the dendrites of some sensory neurons, at the basal pole of the cilium. Presumably the centriole organizes the microtubules which make up the distal cilium, and perhaps the contents of the striated rootlet, where present.
See Centrosome
|
| Centrosome |
|
|
This structure acts as the primary microtubule organizing center (MTOC), and is important both for creation of the mitotic spindle during cell division, and for creation of the cytoplasmic network of microtubules during cytokinesis and during G1 and G2 phases. The centrosome contains two centrioles, plus the intermediate filament protein, IFA, and is generally surrounded by a halo of other fibrous materials including the ZYG-9 protein (Leung et al., 1999). Shortly after fertilization of the early embryo, the centrosome becomes anchored to the plasma membrane asymmetrically near the point of entry of the sperm into the egg; this anchoring point helps to establish the initial polarity of the embryo. Later in cell division, just after duplication of the centrosomes, the two daughter centrosomes show specific movements within the cytoplasm to organize two independent mitotic spindles, including the “rotation” of one centrosome versus its sister, and the “centering” of each centrosome within two different domains to establish separate regions with independently organized axes of polarity.
See Centriole
|
| CEP cell |
CEPDL
CEPDR
CEPVL
CEPVR |
ABplaaaaappa
ABarpapaappa
ABplpaappppa
ABprpaappppa |
All four
are dopaminergic neurons of cephalic sensilla.
See Cephalic sensilla section
|
| Cephalic
nerve cord |
|
|
See Mechanosensory
nerve |
| Cephalic
sensilla |
|
|
Refers specifically to the
sensory specializations of the CEP and CEM neurons and socket and sheath
cells. In some nematode species these sensilla lie on the head (hence
“cephalic”) rather than on the lips (Coomans, 1979), but in C. elegans the cephalic sensilla lie close to the outer labial
(OL) sensilla on the lips proper (See Cephalic sensilla section).
See CEP cell
See CEM cell
|
| Cephalic
sheath cell |
CEPshDL
CEPshDR
CEPshVL
CEPshVR |
ABarpaaaapp
ABarpaaapap
ABplpaaapap
ABprpaaapap |
Refers
to the sheath cells of the cephalic sensilla of the nose. Includes CEPshDL/R,
CEPshVL/R. Their sheet-like processes envelop neuropil of ring and part of ventral ganglion (See Cephalic sensilla section). |
| Cephalic
socket cell |
CEPsoDL
CEPsoDR
CEPsoVL
CEPsoVR |
ABalapapppp
ABalapppppp
ABalppaappp
ABalaapappp |
Socket
cells of the cephalic sensilla (See Cephalic sensilla section). |
| Cephalid |
|
|
A cuticular specialization found in some plant parasitic nematodes, appearing highly refractile, possibly due to thinning of the cuticle within one annulus, local tissue adhesion to the cuticle, or the local passage of a nerve commissure close to the annulus (McLaren, 1976).
See Hemizonid
|
| CEPsh
cell |
|
|
See Cephalic
sheath cell |
| CEPso
cell |
|
|
See Cephalic
socket cell |
| Cervical
papillae |
Anterior
deirid (S) |
|
See Deirid |
| CFP |
|
|
Cyan
fluorescent protein.
See GFP
|
| Channel(s) |
|
|
Narrow
cuticle-lined tubes lying at the three corners of the pharyngeal lumen that
are likely to remain open even when the rest of the lumen is closed during
pharyngeal pumping. It has been proposed that these channels allow outward
flow of fluid from the pharynx during pumping (Albertson
and Thomson, 1976). |
| Cheilorhabdion |
|
|
Specialized
zone of cuticle lining the cheilostom, formed by the cells of hyp1, hyp2
and hyp3. The term refers to this zone as seen by the light microscope. |
| Cheilostom |
Lip cavity
(S) |
|
The most anterior portion of
the buccal capsule or stoma. It is the portion of the anterior digestive tract
which is enclosed by the lips.
See Prostom
See Telostom
|
| Chemical
synapse |
|
|
Area of neuron to neuron communication
where the "presynaptic" process releases the chemical contents
(neurotransmitter or neuropeptide) of a small vesicle into the extracellular
space to signal one or more nearby “postsynaptic” neuron processes.
In C. elegans, the release zone within the presynaptic process
typically shows a small collection of synaptic vesicles and an electron
dense specialization attached to the cytoplasmic face of the plasma membrane.
Unlike higher animals though, the postsynaptic processes rarely display
any membrane density marking the collection of receptors. Chemical synapses
may have one postsynaptic target (monadic), two targets (dyadic) or three
targets (triadic).
See Electrical synapse
See Gap junction
|
| Chemoattractant |
Attractant (S)
Repellant (A) |
|
See Attractant
See Repellant |
| Chemorepellant |
Repellant (S)
Attractant (A) |
|
See Repellant
See Attractant |
| Chemosensation |
|
|
Detection of a chemical cue which elicits a response in the animal so that it can react to food, danger or the presence of other animals. The amphids, which contain 11 pairs of chemosensory neurons, are the primary chemosensory organs. See Bargmann, 2006 for more detail.
See Amphid sensilla section
See Chemotaxis
|
| Chemosensory
ending |
|
|
A specialized,
ciliated portion of a neuron’s dendrite which lies exposed to external
chemical stimuli through a hole in the cuticle. Most chemosensory endings
in C. elegans consist of a neuron and one or more accessory cells
which form a pocket around the cilium to bathe the ending in a fluid-filled
chamber. |
| Chemosensory
nerve |
|
|
A collection
of neuronal processes (dendrites) which run from a discrete group of chemosensory
endings (such as the amphid or phasmid) to their cell bodies and/or their
neuropil. Most chemosensory neurons extend a single dendrite via a chemosensory
nerve to their sensory ending, and a second process (axon) to the neuropil
of a nearby ganglion or nerve ring. |
| Chemotaxis |
|
|
The guided behavior of an animal to approach or avoid the source of a chemical signal in the environment. In C. elegans, chemoreceptors in the lips sense these signals and the animal can be observed to wag its head from side to side, apparently to compare signal strengths and detect the direction of a chemical gradient, before moving its whole body up (positive chemotaxis) or down (negative chemotaxis) the gradient. The animal modulates its rate of turning to redirect itself up or down the gradient (Miller et al., 2005; Bargmann, 2006). Chemotaxis assays have been developed to study this behavior and the cells and genes that are involved in C. elegans. |
| Chemotropism |
|
|
The growth
of a portion of a cell towards or away from a chemical signal. In C. elegans this term has been used in relation to the guidance or repositioning of
muscle arm extensions. |
| Chitinous
layer |
|
|
The most prominent layer within
the eggshell, lying between the inner “lipid layer” (the “inner”
vitelline membrane), and the outer vitelline layer. Although the development
of this layer is still unclear in C. elegans, the chitinous layer
is perhaps secreted by the oocyte itself, just after fertilization.
According to Bird and
Bird (1991), the eggshell is the only place in nematodes where chitin
is known to be utilized. There are protein structures mixed into this
layer in some nematode species as microfibrils or as radial pillars, but
these have not been demonstrated in C. elegans.
See Eggshell
See Oolemma
See Zona pellucida
|
| Chitinous
shell |
|
|
See Eggshell |
| Chord |
|
|
See Cord |
| Chromatin |
|
|
The network of chromosomal material found within the nucleus, made from DNA strands and associated histone proteins. Based upon its staining properties, it is divided into two components, euchromatin (lower staining) and heterochromatin (higher staining). Only the former is thought to be very active, as it is more loosely wound and open to transcriptional machinery. See Schaner and Kelly, 2005. |
| Chromomere |
|
|
A region along the length of a chromosome where the material is more darkly staining due to local condensation of the genetic material. Chromosomal bands consists of local clustering of chromomeres. |
| Chromosome |
|
|
A structure in the cell nucleus which contains the genetic material encoded as DNA and surrounded by histone proteins and other regulatory elements. In C. elegans the normal cell contains 5 pairs of “autosomes” and one or two X chromosomes. |
| Ciliary cap |
Nubbin (S) |
|
Dense material displayed in the cuticle in association with the tip of the sensory cilium as in the cephalic sensory cilia. |
| Ciliary necklace |
|
|
A distinctive arrangement of doublet microtubules that form a circular ring when viewed in cross-section, found within the basal bodies of many chemosensory cilia in C. elegans including the amphids (Perkins et al., 1986). |
| Ciliome |
|
|
A ciliary proteome database which includes cilia-associated proteins (Inglis et al., 2006). |
| Cilium
/ Cilia (/plu) |
|
|
Long, usually motile, whiplike or hairlike appendages that are attached
to eukaryotic cells. Cilia contain an axoneme which is made of a pair
of central microtubules surrounded by nine pairs of microtubules around
the outside (9+2 arrangement). Although the cilia in nematodes are homologous
to the cilia in other animals, they are not motile. To date, no nematode
with motile cilia has been found. Cilia are retained in sensory organs in nematodes in which the specialized sensory ending of a neuron often
features a bundle of microtubules extending distally from a basal body,
most often very distant from the cell soma (For more detail see Inglis et al., 2007).
A wide variety of specializations
are known among the cilia in C. elegans, including local branches,
intracellular dense blobs or dark vesicles, striated rootlets that can
extend proximal to the basal body, and local external attachments to the
basal lamina, the cuticle or within a glial (structural) cell. They are
presumed to be the site of sensory transduction by a specific modality
for which the ciliary specializations are suited; e.g. chemo-, osmo-,
thermo-, mechano-, photo-, etc. Signals transduced by the cilium are carried
to the sensory cell soma by a dendrite and then along an axon to the cell’s
synapses. |
| Circomyarian |
|
|
Describes a muscle cell in which the sarcomeres completely encircle the outer bound of the cell, with the cytoplasmic muscle belly confined to a central core of the cell. Only a few specialized muscles in nematodes may fit this condition.
See Meromyarian
|
| Cistern
/ Cisternae |
|
|
A thin
clear space enclosed by a unit membrane, often occurring as parallel close
fitting spaces. References to cisternae in earlier nematode literature most likely refer to rough ER in a cell soma, or less often to smooth ER
or sarcoplasmic reticulum in distal cell processes. Some cells have distinct
sets of cisternae, including the amphid sheath cells where they are named lamellae, and some of the ray neuron cell bodies of the male tail. |
| Clear
phenotype |
clr |
|
A very
distinctive phenotype in which the adult nematode is remarkably transparent
when viewed by light microscopy due to a single gene mutation, clr-1(Kokel
et al., 1998). These animals also exhibit other morphological changes,
including some bloating and a shorter body length.
clr-1 encodes a receptor tyrosine phosphatase which attenuates signaling by egl-15, a fibroblast growth factor receptor. It appears that these genes act in the hypodermis to regulate fluid homeostasis (Huang and Stern, 2004). |
| Cleidoic |
|
|
Term is used to describe an impermeable eggshell, like that of C. elegans, where there is no exchange of water and fats, and resists physical deformation. Only gases seem to be permeant (Anya, 1976). Such eggs are environmentally isolated and the developing embryo cannot be fed by osmosis through its exterior. |
| Cloaca |
|
|
A structure in the adult male tail, equivalent to the proctodeum (rectum and anus) of the hermaphrodite, where the vas deferens and the intestine open out jointly into an enclosed sinus near the tail tip to discharge their contents. The cloaca refers to the cuticle-lined chamber where these tissues join, including the spicule pouch and ventral opening, but not to the cells themselves, which are all part of the proctodeum (Lints and Hall, 2005).
See Gubernaculum
See Proctodeum
|
| Cloacal
ganglia |
|
|
A pair
of neuronal ganglia in the adult male tail, lying on either side of the
proctodeum, but not found in the hermaphrodite. |
| Clr |
Clear |
|
CLeaR phenotype
or mutation.
See Clear phenotype
|
| Clumping
behavior |
|
|
The tendency
of a population of animals to collect closely together into larger piles
or clumps. This behavior is often observed in normal animals on plates where
the food is about to be exhausted, where they pile up at the edges of the
lawn of bacteria. |
| Coated
pit |
|
|
An intermediate stage of endocytosis,
in which the plasma membrane indents when organized by a coat of clathrin
on the cytoplasmic surface; the coated pit continues to indent and become
more spherical until it breaks off to form a coated vesicle. They have
been seen in neurons near synapses and at the base of sensory cilia and
in coelomocytes (Hall, unpublished). Coated pits are larger in diameter than coated vesicles.
See Caveola / Caveolae
See Coated vesicle
|
| Coated
vesicle |
|
|
A specialized
small vesicle associated with endocytosis, often found at the plasma membrane,
or associated with vesicular trafficking between membranous lamellae deeper
inside a cell, such as at Golgi stacks or rough ER. The outer coat is made
from primarily from clathrin protein. |
| Coelomic
cavity |
|
|
Cavity formed between separate
mesodermal layers which, in vertebrates, includes pleural, peritoneal
and pericardial spaces. The mesodermal layer around the coelomic cavity
is an epithelium proper with a basal lamina on the outside and very often
cilia facing the lumen. There is no true coelomic cavity in the nematodes
but a pseudocoelom.
See Pseudocoelom
|
| Coelomocyte |
|
|
A free-floating
spherical cell lying in the pseudocoelomic cavity of larvae and adult C.
elegans that can endocytose many compounds, possibly for immune surveillance.
There are six coelomocytes in adult hermaphrodites, often lying pairwise
together, and they display prominent cytoplasmic inclusions and vacuoles. |
| Coiler |
|
|
A behavioral phenotype common to some classes of uncoordinated mutations, in which the
animal commonly comes to rest in a tightly coiled posture, forming a spiral,
and tending to remain stationary in this position rather than actively moving
on the plate. In some classes of mutation, the animal may favor coiling
specifically in one handedness, being either a “dorsal coiler”
(with all dorsal muscles in contraction) or a “ventral coiler”
(with all ventral muscles in contraction). Some mutations lead to coiling
when the animal is reversing, but not during forward motion. The mutations
seem to interfere preferentially with neuronal input to the bodywall muscles
so as to block the normal pattern of alternating waves of contraction along
the bodywall which would flex the animal in alternating dorsal and ventral
contraction. |
| Cold
sensitive |
|
|
Referring
to a temperature-sensitive mutation whose phenotype is exacerbated by low
temperature (often 15 or 16oC), but becomes more normal at room temperature. |
| Collar |
|
|
In the spermatozoon, an electron
dense cytoplasmic specialization surrounding an ER membrane through which
the membrane converts to form smaller membranous vesicles on the cis face
of the Golgi apparatus (Wolf
et al., 1978). The two portions of membrane compartment separated
by the collar have also been referred to as the head lobe (smaller) and
body lobe (larger) (Achanzar
and Ward, 1997). After fusion of the membranous organelle with the
spermatozoon’s plasma membrane, the collar is thought to persist
and to reinforce the fusion pore, keeping it open while fibrous contents
from the MO are extruded to the outside surface of the spermatozoon.
See Membranous organelle
|
| Colonizing |
|
|
1) A nematode's ability to invade and reproduce in a local habitat.
2) The ability of a bacteria, fungus or virus to invade the nematode and reproduce there. |
| Comma |
|
|
A middle
stage in embryogenesis of the worm in which the embryo is slightly folded
within the eggshell. |
| Command
interneuron |
Command
neuron (S) |
|
Interneurons whose synaptic
output represent a “final common pathway” to motor neurons.
The synaptic activities of command interneurons lead directly to the control
of locomotion. This group of interneurons include AVA, AVB, AVD, AVE and PVC.
See Interneuron
|
| Commensal |
|
|
A lifestyle in which two species live together (and perhaps to feed together), at no harm to either organism and for the benefit of one (but not both) of the organisms. If both species derive obvious benefits, it is considered mutualism. It has been suggested that C. vulgaris may have a commensal relationship with pillbugs in the wild, living under the shell of the pillbug and perhaps consuming bacteria there (Baird et al., 1994).
See Free living
See Mutualism
See Necromenic
See Parasite
See Phoretic
|
| Commissure |
|
|
The passage of a neuron process
or a bundle of neuron processes between two different nerve cords or between
two bilateral ganglia. In higher animals a commissure always consists
of many processes traveling in parallel to reach a shared destination.
However, in C. elegans, a commissure can consist of a single neuron
process. Such commissural axons in the nematode always travel in direct
contact with a thin coating of the hypodermis, and most run circumferentially
along the outer body wall.
Commissures in C. elegans are:
Amphid commissure: bilateral
Deirid commissure: bilateral
Dorso-rectal commissure: bilateral
Lumbar commissure: bilateral, comprises the full route of axons from preanal
ganglion (ventral side) to dorsal cord
Lumbar-preanal commissure: bilateral, a portion of the lumbar commissure comprising
the ventral to lateral portion only
Motor neuron commissures: each runs either from
left or right side, processes of D type motor neurons that travel
from ventral cord to dorsal cord along the body
See Commissures section
|
| Con |
CONstipated |
|
A defecation
mutant phenotype where animals exhibit delayed or infrequent defecation cycles.
See Defecation motor program
See Constipated/ Constipation
|
| Confocal microscopy |
LSCM (S)
Laser scanning confocal microscopy (S) |
|
This high resolution microscopy is used for imaging fluroescent signals in fixed specimens. |
| Connectivity
diagram |
Wiring
diagram (S) |
|
The overall
pattern by which a set of neurons interacts synaptically with each other. |
| Constipated/Constipation |
|
|
A reduction in the normal rate
of defecation, leading to an abnormal buildup of digestion products inside
the lumen of the hindgut.
See Defecation motor program
|
| Contact
termination |
Branch
termination (S) |
|
A feature in which two neuron
processes touch each other to form a gap junction and terminate at that
point. Very common at the distal extreme of the nerve ring (most commonly
between bilateral processes extending from neurons of the same class)
and along the length of some motor nerves (between functional homologues
such as VD's or DD's). |
| Contralateral |
Ipsilateral (A) |
|
Refers to a cell or cell extension which lies across the midline from a reference object. |
| Continuous
junction |
Smooth
septate junction (S) |
|
An intercellular junction found
only in the spermatheca, in which the two plasma membranes are separated
at a distance (the septa) but show no cytoplasmic density or specialization
to mark the junction. Their presence is most easily noted by MH27 (AJM-1)
antibody staining.
Alternately, this term has been used to characterize extensive adherens
junctions, or “belt desmosomes”, that extend “continuously”
over the entire apposition of two epithelial cells.
See Septate junction
|
| Convulsion |
|
|
A mutant phenotype in which the animal suffers from local contractions or stretching of the bodywall muscles occurring synchronously over all four dorsal and ventral quadrants, so that the local portion of the body seems to lengthen or shorten (Williams et al., 2004). Global changes in length operating on all bodywall muscles synchronously are often called the Rubberband phenotype.
See Rubberband phenotype
|
| Copulatory
apparatus |
Copulatory
structures (S) |
|
The specialized
structures of the male tail which allow the animal to sense the presence
of the hermaphrodite, search her body for the vulval opening, grasp onto
the vulva, and to transfer sperm. |
| Copulatory
bursa |
Bursa
(S) |
|
See Bursa |
| Copulatory
plug |
Mating
plug (S) |
|
Gelatinous
material deposited into the opening of the vulva of the hermaphrodite by
a male at the termination of mating behavior. The plug can produce a visible
swelling that completely covers the vulval region. The plug may preserve
for up to 24 hrs before egg laying forces it to be shed.
Some strains of C. elegans males secrete plug material, while other strains do not
(Barker,
1994). Presence of the plug has been shown to interfere with successful
mating by a second male, and thus protects the sperm from the first male
(the “plugging male”) from competition from sperm from a second
male.
The ability to produce a plug is ascribed to the function of a single
gene, plg-1 (Hodgkin
and Doniach, 1997). The plug material appears to be formed by coagulation
of a secreted yellow liquid, which collects in seminal vesicle during mating,
and is secreted at the end of mating, during which time the male continues
to “linger” near the vulva, after the termination of sperm transfer
(Barker,
1994). plg-1 probably encodes a coagulant, not the yellow liquid
itself, since non-plugging males also exude the yellow liquid. |
| Copulatory
spicule |
|
|
See Spicule |
| Cord |
|
|
This term has multiple, overlapping
uses. It can refer to:
1) The bundled nerves running
along the longitudinal body axis, i.e. “nerve cords”
2) the
hypodermal extensions lying along the sides (dorsal, ventral, left or
right), i.e. “hypodermal cords”
3) the combination of nerve
processes plus hypodermis along the longitudinal body axis
See Hypodermal ridge
See Nerve cord
|
| Core |
Rachis
(S) |
|
The syncytial acellular portion of the germline in the adult male nematode
linking the cell bodies of individual germ cells; it appears completely
the same as the rachis of the hermaphrodite gonad (Wolf
et al., 1978). |
| Corpse |
|
|
This
term can refer either to the dead remains of a whole animal at the end of
life (as in extreme old age, or after deadly insult, or lethal mutation)
or to the dead remains of a cell that has undergone apoptosis or necrosis,
but which has not yet been consumed by engulfment by a neighboring cell. |
| Corpus |
|
|
An extended
anterior region of the pharynx, which is composed of two subdomains, the
procorpus (long and thin) and the metacorpus (first bulb), lying just anterior
to the isthmus and terminal bulb. The muscular corpus lies just posterior
to the pharyngeal epithelium, and anterior to the isthmus. |
| Cortex |
|
|
The outermost
region of the cytoplasm, directly underlying the plasma membrane of a cell.
It is a site of concentration for specific molecular determinants governing
cell development and the point of anchorage for cytoskeletal elements (Munro et al., 2004). The term is often used with regard to the developing blastomere. Other cell
types with distinctive specializations of their cortex include the intestine,
the excretory canal, and the uterus (Gobel et al., 2004).
See Glycocalyx
See Lumen
|
| Cortical
contraction |
|
|
Occurs
in the developing fertilized oocyte prior to first cleavage. These contractions
take place in a polarized fashion relative to the A/P axis of the embryo. |
| Cortical
layer |
Outer layer of cuticle (S)
Epicuticle (S)
Cortical Zone (S) |
|
The outermost layer of the adult cuticle is called the cortical layer, and appears more densely staining
both by light microscopy and by TEM observation. By some accounts the
cortical layer can be subdivided further to identify an external cortical
layer that is the darkest part, an internal cortical layer that is similar
to the matrix layer, and a medial layer.
See Basal
layer
See Fiber layers
See Fibril layer
See Matrix layer
See Epicuticle
|
| Cortical
rearrangement |
|
|
Refers
to changes in anatomical organization in the late stages of primary oocyte
maturation just prior to fertilization, as viewed by light microscopy. |
| Cortical
ruffling |
|
|
Periodic
indentations, or creases, of the first embryonic cell near the time of pseudocleavage
that move laterally along the surfaces of the dividing fertilized oocyte,
as viewed by light microscopy (Hird
and White, 1993). |
| CP
cell |
CP0
CP1
CP2
CP3
CP4
CP5
CP6
CP7
CP8
CP9 |
P2.aap
P3.aapp
P4.aapp
P5.aapp
P6.aapp
P7.aapp
P8.aapp
P9.aapp
P10.aapp
P11.aapp |
Male specific cells in ventral
cord
Male specific cells in ventral cord
Male specific cells in ventral cord
Male specific cells in ventral cord
Male specific motor neurons in ventral cord
Male specific motor neurons in ventral cord
Male specific motor neurons in ventral cord
Male specific motor neurons in ventral cord
Male specific interneurons project into preanal ganglion
Male specific interneurons project into preanal ganglion
|
| Cpa |
|
|
Term
defines CytoPlasmic Appearance alteration mutant or phenotype in early embryo. |
| Crawling |
Gliding (S)
Creeping (S) |
|
The typical undulating locomotory behavior displayed by most nematodes, including C. elegans when moving on a solid or semi-solid substrate (Crofton, 1971; Nicholas, 1975). C. elegans tends to crawl forward more often and more skillfully than backwards, except for adult males, which spend more time moving backwards.
See Swimming
|
| Cristae |
|
|
The internal compartments formed by the inner membrane of a mitochondrion. The foldings of the membrane create a large surface area where the reactions for cellular respiration to take place.
See Mitochondrion
|
| Cross
bridge |
|
|
May refer
to the cell body and lateral branches of the excretory canal process that
connects the left side canal and right side canal into an “H”
shape. The cross bridge also is the site of the secretory membrane at which
the canal cell dumps its contents into the excretory duct and pore. |
| Cross-over |
Decussation
(S) |
|
The crossing
of an axon across the midline is sometimes referred to as "crossing-over"
although the usual term is decussation. |
| Cross
progeny |
|
|
Progeny
that derive from a mating event between male and hermaphrodite parents
and not from self-fertilization within the hermaphrodite, which are termed
self-progeny. |
| Crossing |
|
|
The deliberate production of
cross-progeny by the mating of males and hermaphrodites.
See Backcross
See Outcross
|
| Crumpled
spicules |
|
|
A common deformity in the development
of the spicules of the male tail as a result of certain mutations. This
deformity generally causes male sterility. Whereas normal spicules are
created as long straight rigid structures, supported by a keratinized
outer cuticle, various mutations lead them to become malformed where they
are shorter, bent or too pliable to function in mating behavior.
See Spicule
|
| Cryptobiosis |
|
|
The ability to survive for long periods of time in a metabolically inactive state, whether due to low oxygen, low temperature (including freezing), dessication or even high temperature, among others.
This is different from diapause, but it would seem that larval stage animals are much better suited to survive cryptobiotic conditions than other stages or dauer animals. Similarly, in the laboratory, most mutant strains of C. elegans are best stored for long intervals (years) under liquid nitrogen as freshly starved L1-L2 larvae (Stiernagle, 2006). Some nematode species are much better suited to long term cryptobiotic survival than C. elegans (Nicholas, 1975). |
| Crystalloid |
|
|
A category
of free-floating cytoplasts found in the pseudocoelom of some nematodes
(not known in C. elegans as yet) that accumulate high amounts of
metal sulfides and some protein and carbohydrate into a granular objects. |
| Curl |
Curling (S) |
|
A distinctive posture of the male tail often seen in animals suspended in liquid medium, involving a deep bending on the ventral side. Adult males exposed to high serotonin levels exogenouosly often will curl and sometimes become frozen in this position (Loer and Kenyon, 1993).
|
| Cuticle |
|
|
A rigid external coating which
is secreted by the hypodermis, seam and some interfacial epithelial cells.
It covers the outer body, the major openings into the body cavity (mouth, excretory pore, rectum) and two large sensory bristles, the male spicules. The cuticle of the spicules, the hook, and of some portions of the spicule channels is especially
rigid and is probably sclerotized. Similar sclerotic features are occasionally
noted in the ventral surface of the male tail fan, and are also seen at
the three apices of the buccal cuticle, reinforcing the shape of the cuticle
channels there.
The adult cuticle is comprised of many distinct layers, not all of which
may be evident in any one cross-section by electron microscopy. Larval
and dauer cuticles may look substantially different. The dauer cuticle is relatively much thicker than other stages (Kramer,
1997), as is the cuticle of very old adults (Herndon
et al., 2002). However, neither condition seems to involve the creation
of extra layers, but just the thickening of certain sublayers.
The cuticle of the buccal cavity and pharynx is qualitatively different
from the body cuticle, lacking layers and forming a series of complex
specializations, including the flaps, sieve and grinder. It is virtually
separate from the body cuticle except for tenuous connections through
the “bridging cuticle” (Hall, unpublished).
For more detail see Page and Johnstone, 2007 and WormAtlasHermaphrodite Cuticle and Dauer Cuticle sections.
See Basal
layer
See Boundary
zone
See Channel
See Cortical layer
See Fibril layer
See Molt
See Matrix layer
|
| Cuticle
collagens |
|
|
Those collagen proteins that are expressed in the nematode cuticle at
some time during the animal’s development or adult life stages. Genes
encoding collagen proteins are among the larger gene families within the C. elegans genome. Some collagens are not expressed in cuticle, but
instead help to form the basal laminae of various tissues (for more detail see Page and Johnstone, 2007). |
| Cuticle
flaps |
|
|
Three
flaps extend inward from the lining of the buccal cuticle to restrict flow
at the rear of the buccal cavity (Wright
and Thomson, 1981). These flaps extend from the pharyngeal muscle cells
m1 and m2. They are made of specialized cuticle and appear very electron
dense in thin section, suggesting a sclerotic hardening to stiffen the flaps
and the entryway to the true pharynx. |
| Cuticle
furrows |
|
|
Shallow circumferentially-oriented indentations in the cuticle. The ridges between furrows are the annuli.
See Annuli
|
| Cuticular
nubbin |
CN |
|
See Nubbin |
| Cyk |
Cytokinesis abnormal (S) |
|
CYtoKinesis
defect. Term defines abnormality in the division of a cell into two daughter
cells
following mitosis. |
| Cytoneme |
|
|
Thin
tapered processes extended by somatic sheath cell #1 over the surface of
the unsheathed gonad, near the transition zone (Hall
et al., 1999; Hubbard
and Greenstein, 2000). |
| Cytoplast |
Minicell
(S) |
|
A non-viable cell fragment
which has broken off from a parent cell, or also it can be a residual fragment of
a dead or dying cell. Cytoplasts most often contain no nuclear material,
and thus cannot reproduce or maintain themselves (Sulston
and Horvitz, 1981). Other categories of cytoplasts may exist as exudates
or waste products which assemble spontaneously outside of any cell, such
as crystalloids in some species.
See Bleb
See Brush
See Crystalloid
|
| Cytosome |
|
|
Archaic term for an organelle lying within a cell’s cytoplasm which might be either an endosome (membrane-bound) or a free granule, possibly comprised of yolk or lipid (Popham and Webster, 1979). |


{kind=link}