|
The Embryonic Cell Lineage of the Nematode Caenorhabditis elegans
J.E. Sulston*, E. Schierenberg†, J.G. White* and J.N. Thomson*
*M.R.C. Laboratory of Molecular Biology, Hills Road, Cambridge, CB2 2QH, England
and
†Department of Molecular Biology, Max-Planck Institute for Experimental Medicine, 3400 Gottingen Federal Republic of Germany
Developmental Biology (1983) 100: 64-119
doi: 10.1016/0012-1606(83)90201-4
(Received November 3, 1982; accepted in revised form January 14, 1983)
Nomenclature note for this paper: MS was formerly MSt, U was E, Y was C, W was P0.a, QL was Q1, and QR was Q2.
Abstract - Introduction -
Material & Methods -
Results -
Discussion -
Acknowledgments
- References
WA editors' note: In this paper, the positions of the descendants of Caaa and Cpaa are reversed in Figures 7a (260 min), 10 (290 min), 13 (L1) and 14 (L1). At 260 min, the four nuclei descended from Caaa are to be found on the left of the dorsal midline and those from Cpaa on the right. At about 310 min of embryogenesis, most of the dorsal hypodermal nuclei migrate across the midline to reside on the side opposite to that of their origin (Fig. 10). Thus, after this stage, the four descendants of Cpaa reside on the left side and Caaa on the right side (Figs. 13 and 14a). Note that Fig. 5 is correct with Caa to the left (and anterior) of Cpa (Loer C. et al, WBG 10(3):120).
Abstract
The embryonic cell lineage of Caenorhabditis elegans has been traced from zygote to newly hatched larva, with the result that the entire cell lineage of this organism is now known. During embryogenesis 671 cells are generated; in the hermaphrodite 113 of these (in the male 111) undergo programmed death and the remainder either differentiate terminally or become postembryonic blast cells. The embryonic lineage is highly invariant, as are the fates of the cells to which it gives rise. In spite of the fixed relationship between cell ancestry and cell fate, the correlation between them lacks much obvious pattern. Thus, although most neurons arise from the embryonic ectoderm, some are produced by the mesoderm and a few are sisters to muscles; again, lineal boundaries do not necessarily coincide with functional boundaries. Nevertheless, cell ablation experiments (as well as previous cell isolation experiments) demonstrate substantial cell autonomy in at least some sections of embryogenesis. We conclude that the cell lineage itself, complex as it is, plays an important role in determining cell fate. We discuss the origin of the repeat units (partial segments) in the body wall, the generation of the various orders of symmetry, the analysis of the lineage in terms of sublineages, and evolutionary implications.
Contents. Introduction. Materials, methods, and background information. Culture. General biology. Light microscopy. Electron microscopy. Strategy of observation. Reliability. Nomenclature. Results and comments. General description. Invariance. Cell divisions and cell movement. The founder cells. Gastrulation. Later cell movements. Migrations. Programmed cell death. Other nematode species. Tissue description. Hypodermis. Excretory system. Nervous system. Mesoderm (excluding the pharynx). Alimentary tract. Gonad. Cell interaction experiments. Early ablations. Body muscle from C and D. MS lineage. AB lineage. Other late ablations. Summary. Discussion. Invariance, cell autonomy, and cell interaction. Embryonic germ layers and cell fate. Lineal boundaries and functional boundaries. Segments. Sublineages. Programmed cell death and sexual dimorphism. Rotational symmetry. Symmetry and asymmetry. Conclusion.
Introduction
This report marks the completion of a project begun over one hundred years ago -namely the determination of the entire cell lineage of a nematode. Nematode embryos were attractive to nineteenth century biologists because of their simplicity and the reproducibility of their development, and considerable progress was made in determining their lineages by the use of fixed specimens (reviewed by Chitwood and Chitwood, 1974). By the technique of Nomarski microscopy, which is nondestructive and yet provides high resolution, cells can now be followed in living larvae (Sulston and Horvitz, 1977; Kimble and Hirsh, 1979; Sternberg and Horvitz, 1981) and eggs (Deppe et al, 1978; this paper). The use of living material lends a previously unattainable continuity and certainty to the observations, and has permitted the origin and fate of every cell in one nematode species to be determined. Thus, not only are the broad relationships between tissues now known unambiguously but also the detailed pattern of cell fates is clearly revealed.
Our current interest in the cell lineage of a particular nematode, Caenorhabditis elegans, has arisen as part of a larger research effort comprising genetic, anatomical, and biochemical approaches to the development of this animal. The lineage is of significance both for what it can tell us immediately about relationships between cells and also as a framework into which future observations can be fitted.
The main purpose of this article is to present the embryonic cell lineage. A brief description of morphogenesis is given, although this is not intended to be an exhaustive account of cell movements and interactions. A few cell ablation experiments are described; these results are limited in scope, but do give some indication of the developmental flexibility (or lack of it) in the system. In addition, a key to differentiated cell types is provided as an Appendix.
Materials, Methods and Background Information
Culture
General Biology of C. elegans
Light Microscopy
Electron Microscopy
Strategy of Observation
Reliability
Nomenclature
Culture
Caenorhabditis elegans (Bristol), strain N2, was maintained according to Brenner (1974). Turbatrix aceti and Aphelencoides blastophthorus were kindly given to us by David Hooper, Rothamsted Experimental Station, Harpenden, Herts, England; Panagrellus redivivus was obtained from Rothamsted in 1976, and is the strain studied by Sternberg and Horvitz (1981, 1982). T. aceti and P. redivivus were maintained in the same way as C. elegans; A. blastophthorus was grown on NGM plates infected with mixed fungi.
General Biology of C. elegans
C. elegans is a free-living nematode which has two sexual forms: a self-fertilising hermaphrodite and a male. Development begins in the egg, and continues through four larval stages (L1-L4) to the adult. A newly hatched L1 is depicted in Fig. 1. The general anatomy of the newly hatched L1 larva has been described by Sulston and Horvitz (1977). As far as it goes this account is accurate except for the precise cell count in the head and the omission of the intestino-rectal valve (virL and virR, see Figs. 1 and 8c). Postembryonic cellular development has been described for the gonad by Kimble and Hirsh (1979), for the male tail by Sulston et al. (1980), and for other systems by Sulston and Horvitz (1977). Other references will be found in the tissue-by-tissue description below.
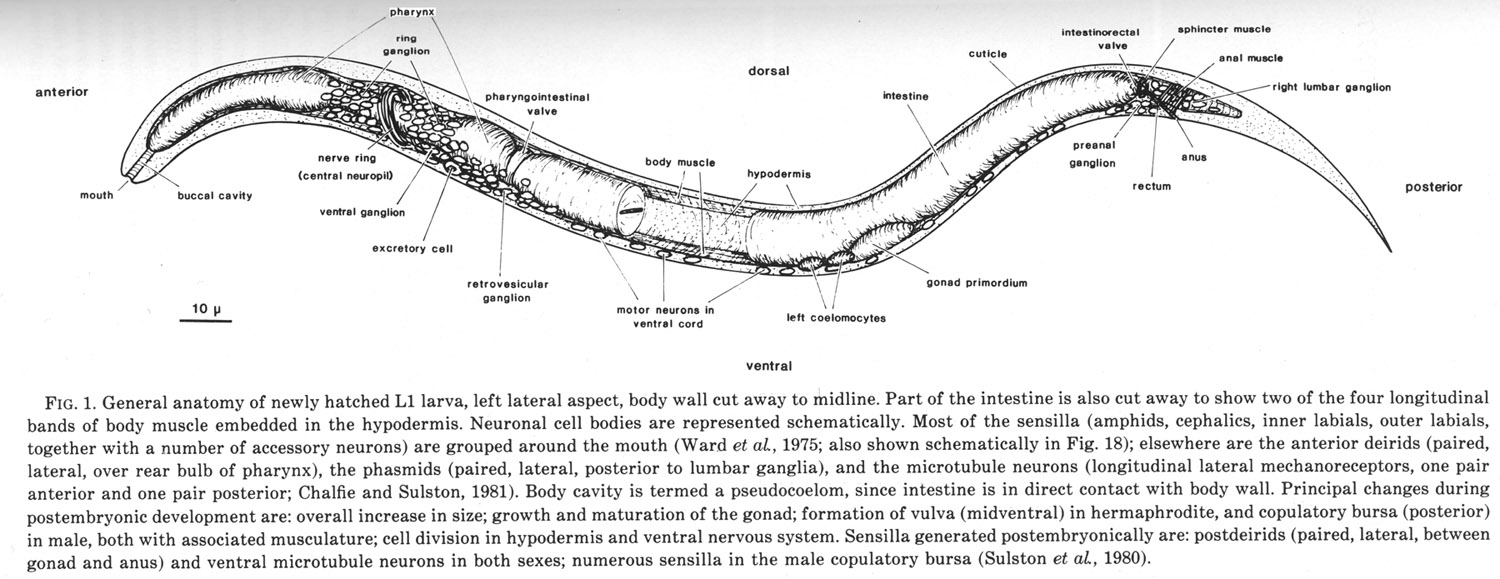
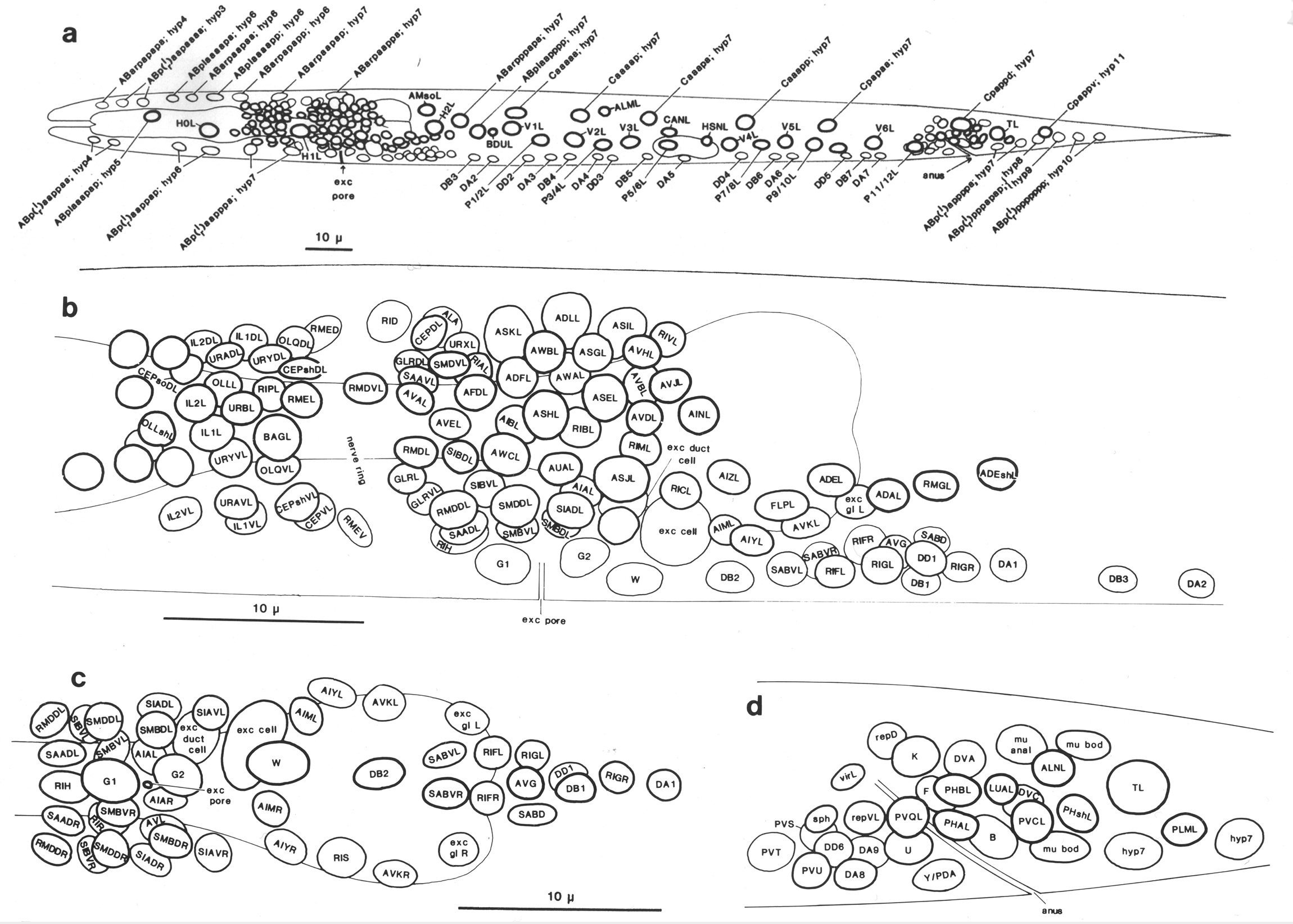
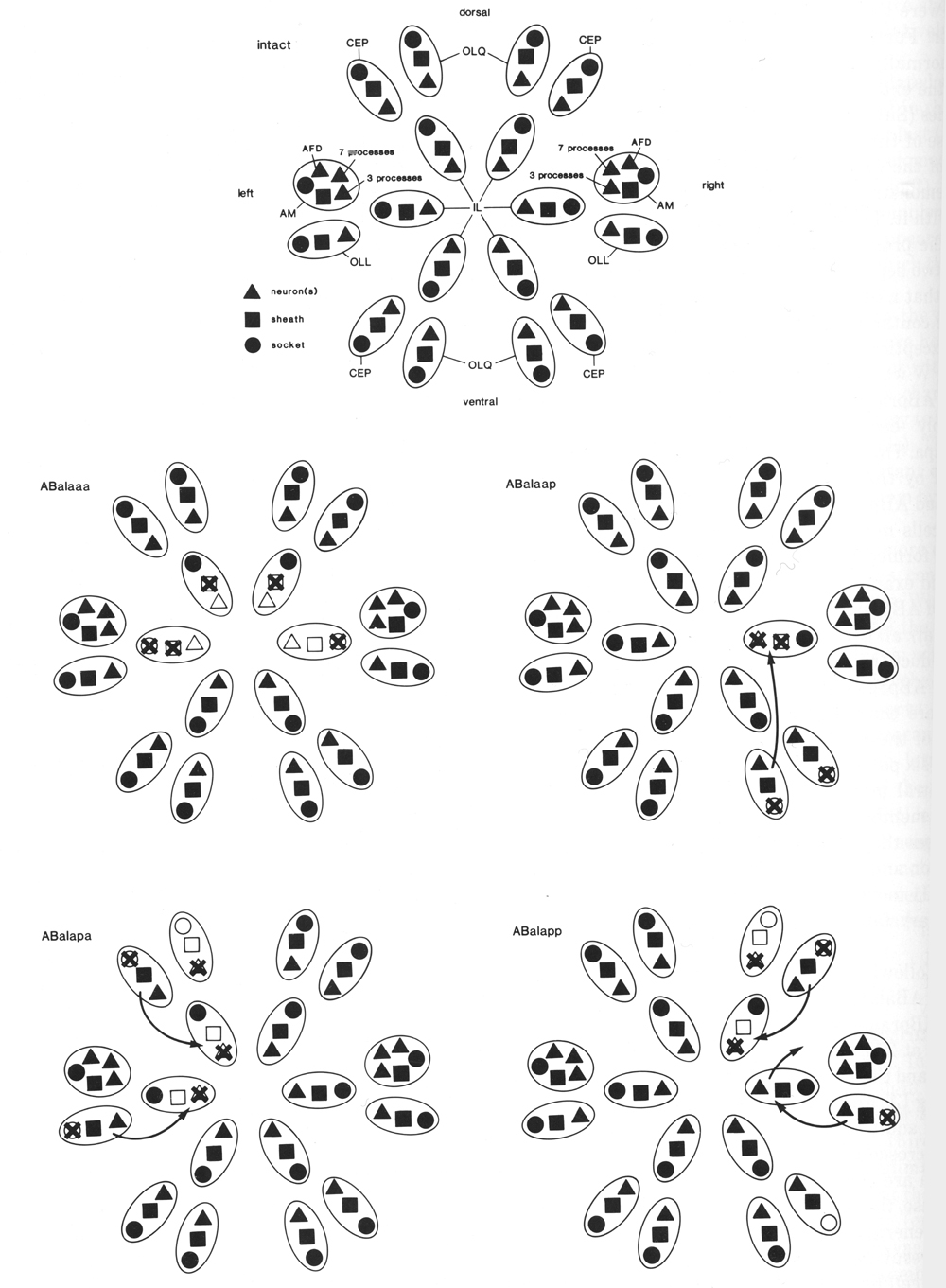
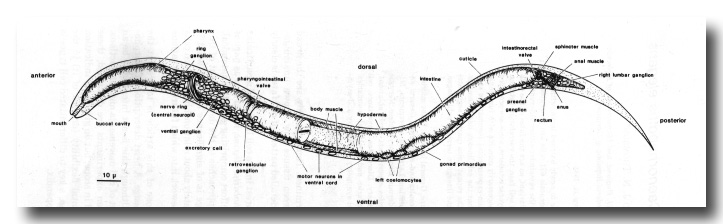 Figure 1. General anatomy of newly hatched L1 larva, left lateral aspect, body wall cut away to midline.
Part of the intestine is also cut away to show two of the four longitudinal bands of body muscle embedded in the hypodermis. Neuronal cell bodies are represented schematically. Most of the sensilla (amphids, cephalics, inner labials, outer labials, together with a number of accessory neurons) are grouped around the mouth (Ward et al, 1975; also shown schematically in Fig. 18); elsewhere are the anterior deirids (paired, lateral, over rear bulb of pharynx), the phasmids (paired, lateral, posterior to lumbar ganglia), and the microtubule neurons (longitudinal lateral mechanoreceptors, one pair anterior and one pair posterior; Chalfie and Sulston, 1981). Body cavity is termed a pseudocoelom, since intestine is in direct contact with body wall. Principal changes during postembryonic development are: overall increase in size; growth and maturation of the gonad; formation of vulva (midventral) in hermaphrodite, and copulatory bursa (posterior) in male, both with associated musculature; cell division in hypodermis and ventral nervous system. Sensilla generated postembryonically are: postdeirids (paired, lateral, between gonad and anus) and ventral microtubule neurons in both sexes; numerous sensilla in the male copulatory bursa (Sulston et al, 1980).
Figure 1. General anatomy of newly hatched L1 larva, left lateral aspect, body wall cut away to midline.
Part of the intestine is also cut away to show two of the four longitudinal bands of body muscle embedded in the hypodermis. Neuronal cell bodies are represented schematically. Most of the sensilla (amphids, cephalics, inner labials, outer labials, together with a number of accessory neurons) are grouped around the mouth (Ward et al, 1975; also shown schematically in Fig. 18); elsewhere are the anterior deirids (paired, lateral, over rear bulb of pharynx), the phasmids (paired, lateral, posterior to lumbar ganglia), and the microtubule neurons (longitudinal lateral mechanoreceptors, one pair anterior and one pair posterior; Chalfie and Sulston, 1981). Body cavity is termed a pseudocoelom, since intestine is in direct contact with body wall. Principal changes during postembryonic development are: overall increase in size; growth and maturation of the gonad; formation of vulva (midventral) in hermaphrodite, and copulatory bursa (posterior) in male, both with associated musculature; cell division in hypodermis and ventral nervous system. Sensilla generated postembryonically are: postdeirids (paired, lateral, between gonad and anus) and ventral microtubule neurons in both sexes; numerous sensilla in the male copulatory bursa (Sulston et al, 1980).
Light Microscopy
C. elegans eggs were transferred from plates which contained ample bacteria to water in a watch glass; alternatively, young eggs were obtained by cutting gravid hermaphrodites in water. With the help of a 50X dissecting microscope about 50 eggs of approximately the required age were selected; they were transferred to a layer of 5% agar, any that were in contact were moved apart with a fine hair, and they were viewed by Nomarski optics (Sulston and Horvitz, 1977; Sulston et al, 1980). A single egg which happened to be at the required stage and in an appropriate orientation was chosen for observation. Representative specimens are shown in Fig. 2.
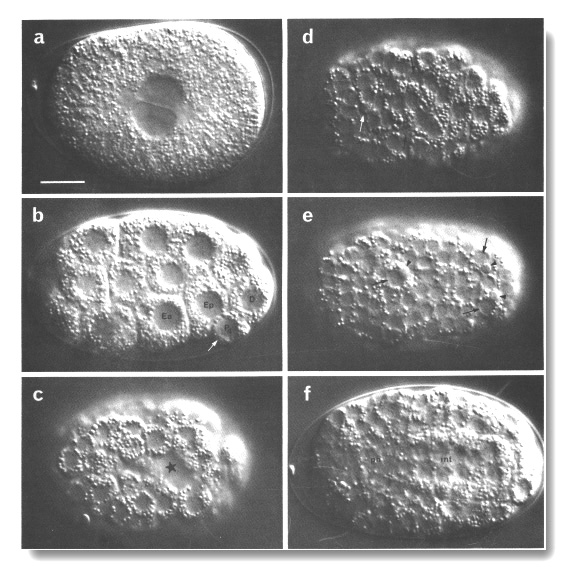
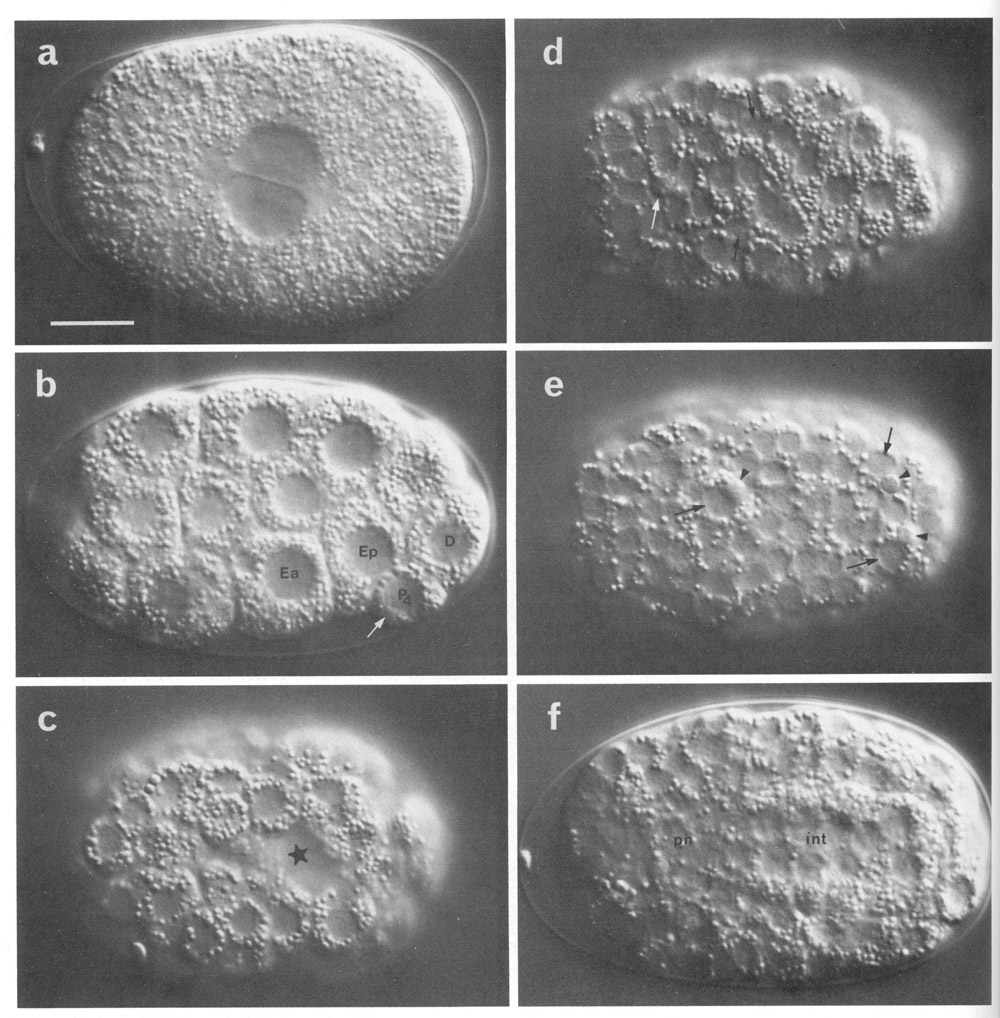
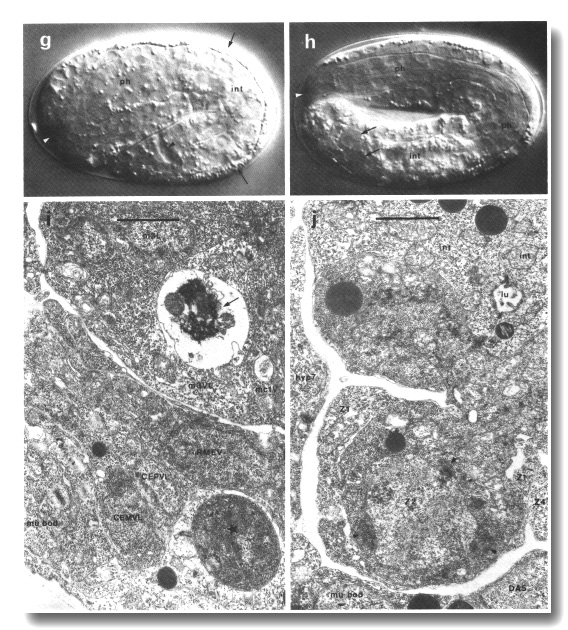 Figure 2. Photomicrographs of embryos, (a-h) Nomarski optics, Microflash, anterior to left. Bar = 10 µm. A few landmark features are marked on the photographs, but no attempt has been made to label all the cells; for reliable identifications the line drawings should be used, preferably in conjunction with at least some tracing of the lineage. (a) Just before first cleavage. Midplane. Male and female pronuclei are apposed; first polar body visible beneath eggshell at anterior end (where it typically but not invariably resides). (b) Beginning of gastrulation. Left lateral aspect, midplane; cf. Fig 5. Ea and Ep are moving dorsally, into the interior. P4 is recognisable by its small size, by the germinal plasm or nuage (arrowed; Strome and Wood, 1982; Krieg et al, 1978) around its nucleus, and by the distinctness of its nuclear membrane. (c) Late gastrulation (ca. 210 min). Ventral aspect, superficial plane; cf. Fig. 6. The cleft through which the MS cells have just entered is starred. (d) ca. 280 min. Dorsal aspect, superficial plane; cf. Fig. 7a. Small neuroblasts anteriorly; larger hypodermal cells, loaded with granules, posteriorly. Furrows can be seen between hypodermal cells (cf. Fig. 10). White arrow, dying ABarpaaapp; black arrows, ADEshL and ADEshR. (e) ca. 260 min. Ventral aspect, superficial plane; cf. Fig. 7b. Small neuroblasts over entire surface. Dying cells (arrowheads) are engulfed by their sisters (arrowed): ABplpappaa, ABplpppapa, ABprpppapa. (f) ca. 280 min. Dorsal aspect, midplane. int, cylinder of intestinal cells, nine nuclei in this focal plane, cytoplasm heavily loaded with granules; ph, cylinder of pharyngeal cells, less distinctive, contain few granules, (g) First movement (ca. 430 min). Left lateral aspect, midplane; cf. Fig. 8c. int, intestine; ph, pharynx; white arrowhead, anterior sensory depression; black arrowhead, rectum; arrows, dorsal hypodermal ridge, heavily loaded with granules, (h) Threefold, rolling. Only the anterior two-thirds of the embryo is in focus. White arrowhead points to mouth, linked by pharynx (ph) to intestine (int). Arrows point to germ cells. (i) ca. 430 min; electron micrograph of transverse section to show phagocytosed cells. Recent death (starred) lies within a ventral hypodermal cell; older death (arrowed) lies within a pharyngeal muscle. Pharyngeal lumen seen at upper right, surrounded by desmosomes between muscles and marginal cells. Outer surface of embryo, seen at lower left, is covered by a tenuous membrane in addition to the hypodermal basement membrane. Bar = 1 µm. (j) ca. 470 min; electron micrograph of transverse section, to show protrusion of lobes from germ cell (Z2) into two intestinal cells (int). Germ cells are united via lobes, but Z3 is not visible in this section. Germinal plasm or nuage (arrowheads; cf. Pig. 2b) visible around Z2 nucleus. Lumen (lu) of intestine is sealed by desmosomes; its wall carries microvilli. Bar = 1 µm.
Figure 2. Photomicrographs of embryos, (a-h) Nomarski optics, Microflash, anterior to left. Bar = 10 µm. A few landmark features are marked on the photographs, but no attempt has been made to label all the cells; for reliable identifications the line drawings should be used, preferably in conjunction with at least some tracing of the lineage. (a) Just before first cleavage. Midplane. Male and female pronuclei are apposed; first polar body visible beneath eggshell at anterior end (where it typically but not invariably resides). (b) Beginning of gastrulation. Left lateral aspect, midplane; cf. Fig 5. Ea and Ep are moving dorsally, into the interior. P4 is recognisable by its small size, by the germinal plasm or nuage (arrowed; Strome and Wood, 1982; Krieg et al, 1978) around its nucleus, and by the distinctness of its nuclear membrane. (c) Late gastrulation (ca. 210 min). Ventral aspect, superficial plane; cf. Fig. 6. The cleft through which the MS cells have just entered is starred. (d) ca. 280 min. Dorsal aspect, superficial plane; cf. Fig. 7a. Small neuroblasts anteriorly; larger hypodermal cells, loaded with granules, posteriorly. Furrows can be seen between hypodermal cells (cf. Fig. 10). White arrow, dying ABarpaaapp; black arrows, ADEshL and ADEshR. (e) ca. 260 min. Ventral aspect, superficial plane; cf. Fig. 7b. Small neuroblasts over entire surface. Dying cells (arrowheads) are engulfed by their sisters (arrowed): ABplpappaa, ABplpppapa, ABprpppapa. (f) ca. 280 min. Dorsal aspect, midplane. int, cylinder of intestinal cells, nine nuclei in this focal plane, cytoplasm heavily loaded with granules; ph, cylinder of pharyngeal cells, less distinctive, contain few granules, (g) First movement (ca. 430 min). Left lateral aspect, midplane; cf. Fig. 8c. int, intestine; ph, pharynx; white arrowhead, anterior sensory depression; black arrowhead, rectum; arrows, dorsal hypodermal ridge, heavily loaded with granules, (h) Threefold, rolling. Only the anterior two-thirds of the embryo is in focus. White arrowhead points to mouth, linked by pharynx (ph) to intestine (int). Arrows point to germ cells. (i) ca. 430 min; electron micrograph of transverse section to show phagocytosed cells. Recent death (starred) lies within a ventral hypodermal cell; older death (arrowed) lies within a pharyngeal muscle. Pharyngeal lumen seen at upper right, surrounded by desmosomes between muscles and marginal cells. Outer surface of embryo, seen at lower left, is covered by a tenuous membrane in addition to the hypodermal basement membrane. Bar = 1 µm. (j) ca. 470 min; electron micrograph of transverse section, to show protrusion of lobes from germ cell (Z2) into two intestinal cells (int). Germ cells are united via lobes, but Z3 is not visible in this section. Germinal plasm or nuage (arrowheads; cf. Pig. 2b) visible around Z2 nucleus. Lumen (lu) of intestine is sealed by desmosomes; its wall carries microvilli. Bar = 1 µm.
T. aceti eggs normally develop within the parent; they are both mechanically and osmotically fragile, and were therefore mounted on 1% agar in isotonic medium. Agar was dissolved in boiling water to a concentration of 2%, cooled to about 50°C, and mixed with an equal volume of a solution containing 180 mM NaCl, 8 mM KCl, 36 mM CaCl2, 36 mM MgS04, 10 mM Hepes, pH 7.2 (cf Laufer et al, 1980). A single gravid adult was cut in half in water, and the extruded eggs were immediately pipetted onto a prepared agar layer. Even with these precautions, development continued for only about 7 hr, representing one-quarter of embryogenesis in T. aceti. However, the successive cell patterns were reproducible (and also similar to those of C. elegans), so the lineage could be traced unambiguously as described below (Strategy of Observation).
P. redivivus eggs were mounted like those of T. aceti, except that the gravid adults were placed directly on the agar layer and cut in half with scissors; the eggs were gently moved away from their parents with a fine hair.
A. blastophthorus eggs were teased out from agar/ fungal blocks into water, and mounted like those of C. elegans.
Electron Microscopy
The egg shell excludes the usual fixatives and embedding media, and must be made permeable in some way before conventional methods can be applied.
The eggs examined in the course of the present work were first treated with NaOCl (2% available chlorine, 5-10 min); they were then fixed with OsO4 (1%, 1 hr) and embedded and sectioned in the usual way (Ward et al, 1975). In an improved version of this method the egg is treated with chitinase or l-phenoxy-2-propanol after the NaOCl, and then prefixed with glutaraldehyde (Albertson and Thomson, 1982).
Permeabilisation with hypochlorite is always rather erratic; a better method for dealing with an individual egg is to puncture the shell with a laser microbeam (Schierenberg and Cole, in von Ehrenstein et al, 1981).
For reconstruction of the anterior sensilla, L4 larvae and adults were prepared as follows. The nematode was transferred to 3% glutaraldehyde in 0.1 M Hepes, pH 7.4, and immediately cut in the posterior half. After about 2 min a second cut was made in the anterior half; after 1 hr the head was washed three times with 0.1 M Hepes, pH 7.4, and then postfixed with 1% OsO4 in the same buffer for 1 hr. The specimen was embedded as usual (Ward et al, 1975), and about 200 serial sections were cut from the anterior tip.
Strategy of Observation
All of the embryonic lineage was followed by direct observation. This method currently gives the best resolution in space and time, but has the disadvantage that the number of cells which can be followed in a single individual is limited by the short-term memory of the observer. Events were recorded by sketching the nuclei, using a colour code to indicate depth. A camera lucida was used at first, but the effective resolution was reduced by this accessory, and the additional illumination required tended to damage the specimen. The best aid proved to be a pair of gossamer cross hairs in one eyepiece, under which the nucleus of interest could be located with the help of a gliding stage. The light was blocked whenever the specimen was not being viewed.
The earlier part of the lineage was also analysed using videotape recordings, and much of this work has already been described (Deppe et al, 1978). The advantages of this technique are considerable: a permanent record is created, in which the cell lineage can be followed at leisure and in which long-range comparisons of cell movements and the timing of events can be made. However, it was not possible to trace the later cell divisions, particularly those taking place in the interior of the embryo, in this way. In order to resolve a small dividing nucleus it is necessary to be able to focus through it without jitter or excessive electronic noise, and at frequent intervals; this is not easily achieved with current equipment. Nevertheless, videotaping remains the technique of choice for studying early embryogenesis both in the wild type and in mutants and experimental animals.
Fortunately for the direct observer, it is unnecessary to follow the lineage from the beginning for each terminal cell. The developing egg displays a succession of reproducible patterns, in which previously identified cells can be recognised. Some of the more useful ones are shown in Figs. 5-8. Although these diagrams were prepared with the aid of a camera lucida they are not intended to show the absolute positions of cells, which in any case vary appreciably from one individual to another; what is reproducible is the neighbourhood of each cell at a given time. The patterns change rapidly, but the behaviour of each cell is characteristic and provides an additional check on its identity. An inexperienced observer should be able to identify nuclei in the diagrams unambiguously by starting one division earlier and checking the arrangement of sister cells.
When placed on an agar layer under a coverslip, the embryo adopts a predictable, though age-dependent, orientation. At the four-cell stage it turns to display either the left or the right side; at gastrulation (100-150 min) it turns from left to dorsal or from right to ventral (these turns are only about 45°, because of the arrangement of blast cells); finally, at 350-400 min, the growing tail forces a return to a lateral aspect. No means of constraint compatible with good resolution was found to prevent these turns, but they were controlled by selection of obliquely oriented embryos at appropriate times.
Cell deaths were recognised by a characteristic increase in refractility, followed by shrinkage and disappearance (Sulston and Horvitz, 1977; Robertson and Thomson, 1982).
The final step in our analysis was to identify the surviving cells in terms of the known larval and adult anatomy. As many cells as possible were identified at 430 min (by comparison with serial section reconstructions of animals at this stage), because thereafter observation is much more difficult on account of movement of the embryo. The most reliable assignments at this time are for cells which have already formed desmosomes or other structural connections (i.e., hypodermis, body muscles, sensory nervous system, alimentary tract) but some useful clues to the identity of other neurons can be gleaned from the initial outgrowth of their processes.
The cells which were not identifiable at this stage (mainly interneurons and motorneurons) were followed in small groups until the animal hatched. These observations are tedious because from 450 min onwards the embryo rotates continually about its longitudinal axis, and it is necessary to train the eye to rapid pattern recognition for each cell group in turn. After hatching, some cells were identified from previously known L1 anatomy (J. G. White et al, unpublished) and the remainder were traced into the adult. This is a relatively easy task because the larvae do not rotate and the patterns change only slowly, so that an entire ganglion can be followed in one individual.
Reliability
The lineage is based on a minimum of two direct observations for unique events, and one direct observation on each side for bilaterally symmetrical events. However, the great majority of events received considerably more confirmation than this. In addition, the earlier part of the lineage, extending to the terminal divisions in the intestine and the lateral hypodermis, was followed independently by videorecording.
Perhaps it is too much to hope that we have entirely avoided errors. It is worth emphasising, however, that any which have arisen are clerical in nature; there are no guesses, and the validity and reproducibility of a particular event can readily be checked by further observation.
Nomenclature
The system of lineage nomenclature is essentially that described by Sulston and Horvitz (1977). Certain key blast cells are given arbitrary names comprising uppercase letters and numbers; their progeny are named by adding lowercase letters indicating the approximate division axis according to an orthogonal coordinate system (a, anterior; p, posterior; l, left; r, right; d, dorsal; v, ventral); the next generation of cells is named by appending further letters in the same way; and so on. Existing blast cell names have been retained as far as possible, but certain changes are desirable to avoid confusion whilst conforming to our system: MS was formerly MSt, U was E, Y was C, W was "P0.a," QL was Q1, and QR was Q2. It should be noted that AB and B are entirely separate names, as are P0-P4 and P1-P12. A pair of cells may be designated by the use of internal parentheses, e.g., MS(a/p)pa means MSapa and MSppa. Pairs of identical postembryonic cells, lying on the left and right sides of the animal, have previously been given identical names; they are now distinguished by the addition of symmetry operators, as defined in the next paragraph. Sometimes developmental stages are named after the number of progeny generated by a particular founder cell; e.g., MS8 means that MS has divided into eight cells.
Functional names are listed alphabetically in the Appendix, and follow a variety of systems:
Neurons and supporting cells other than those in the pharynx: White et al, (in preparation). This system is largely self-explanatory (see Appendix), but note that it uses symmetry operators as suffixes to distinguish cells which differ only in position (A, anterior; P, posterior; D, dorsal; V, ventral; L, left; R, right).
Pharynx: Albertson and Thomson (1976), with the addition of symmetry operators.
Hypodermis: See Figs. 12, 13, 14; symmetry operators are used for the arcades and hypodermal rings 1 and 2.
Muscle: see Fig. 15.
Gonad: Kimble and Hirsh (1979).
Intestine: see Fig. 8c.
In descriptions of cell division the terms "equal" and "unequal" refer to the relative sizes of the daughters; "symmetrical" means that the daughters not only are equal in size but also are (or subsequently become) disposed symmetrically across the midline; "equational" means that the daughters are supposedly of equal developmental potential.
Results and Comments
General Description
Invariance
Cell Divisions and Cell Movement
The Founder Cells
Gastrulation
Later Cell Movements
Migrations
Programmed Cell Death
Other Nematode Species
Tissue Description
Hypodermis
Excretory System
Nervous System
Mesoderm (Excluding the Pharynx)
Alimentary Tract
Buccal cavity, pharynx and pharyngo-intestinal valve
Intestine
Intestino-rectal valve and rectum
Gonad
Cell Interaction Experiments
Early Ablations
Body Muscle from C and D
MS Lineage
AB Lineage
Other late ablations
Summary
GENERAL DESCRIPTION
Invariance
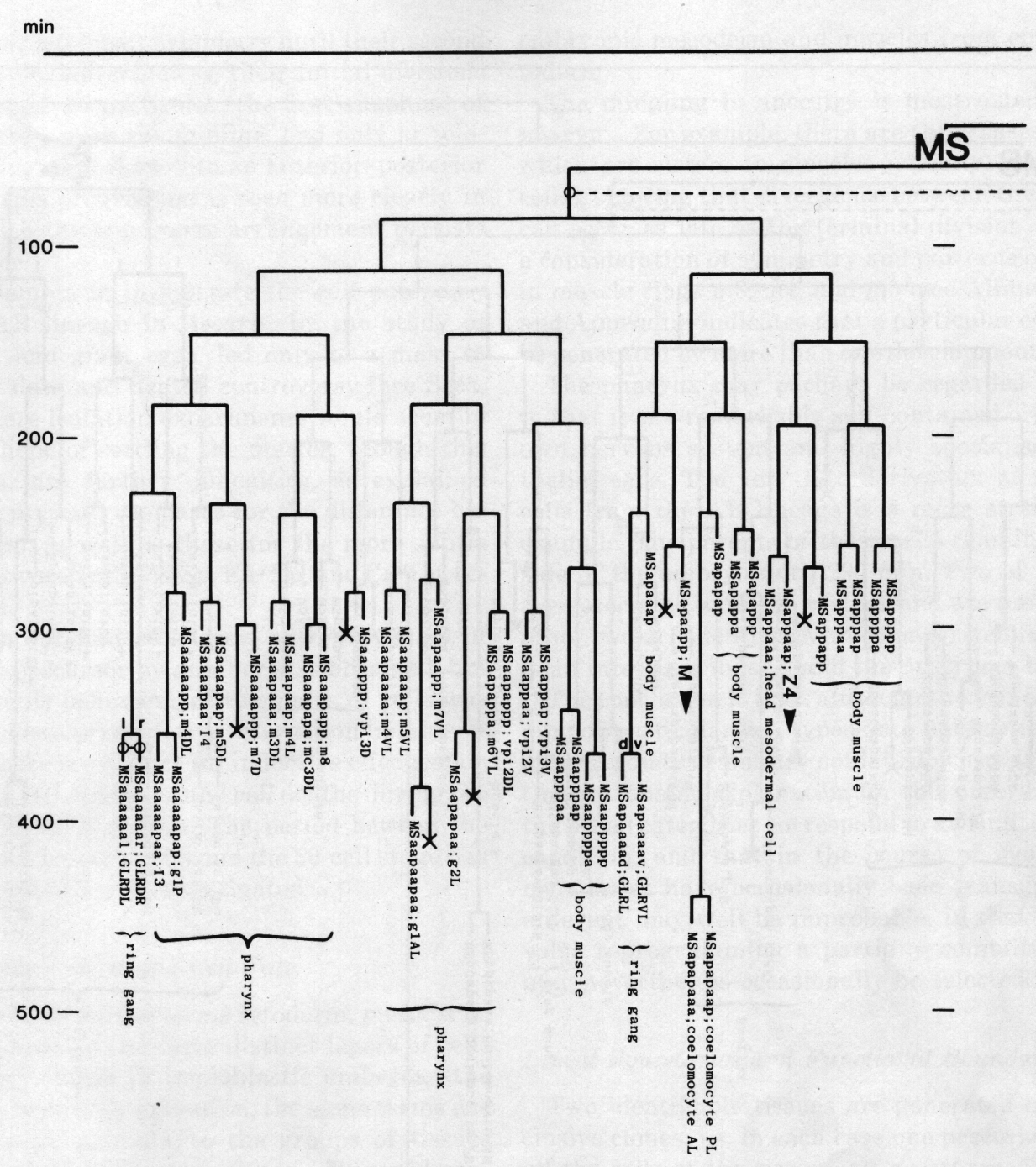
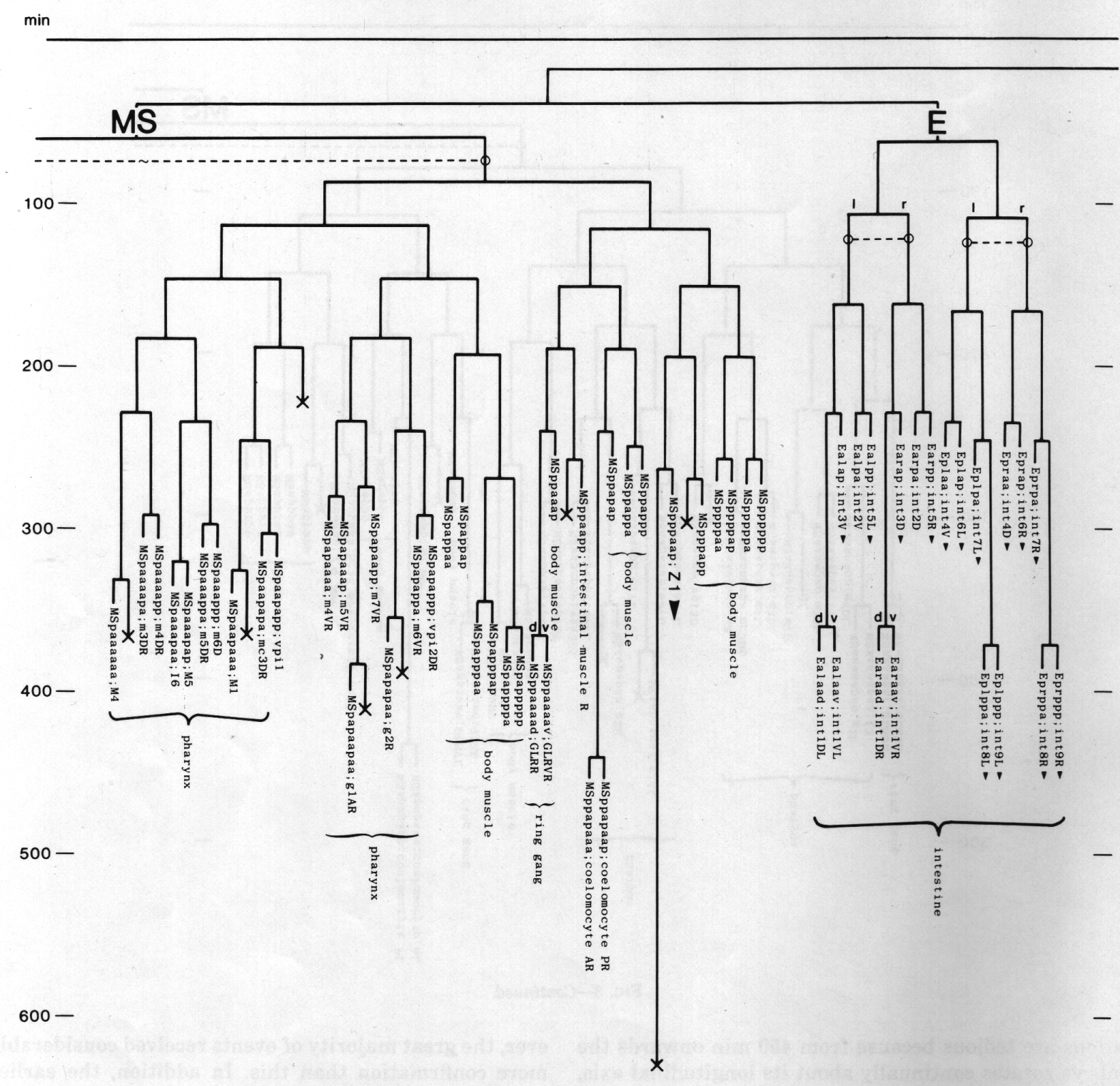
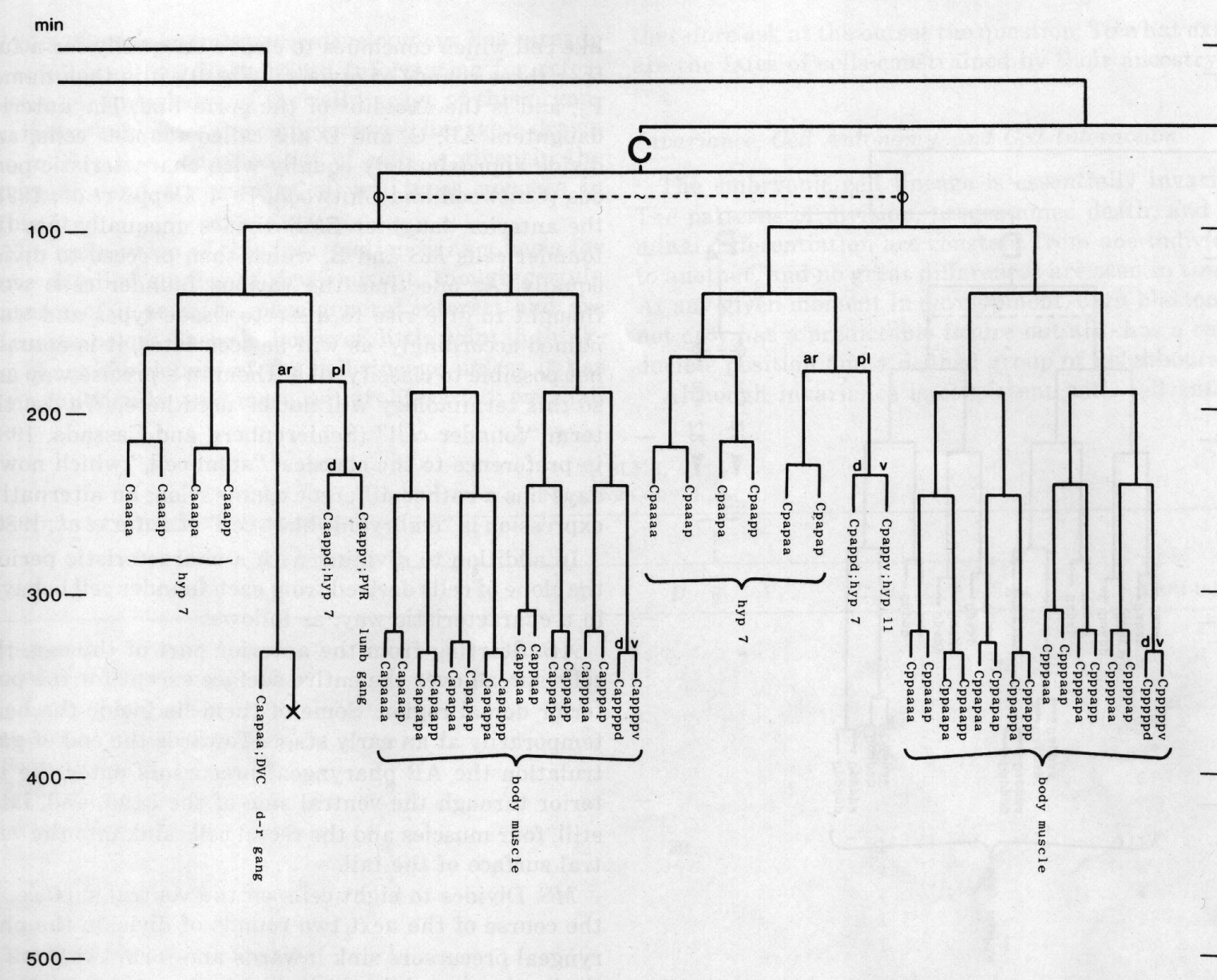
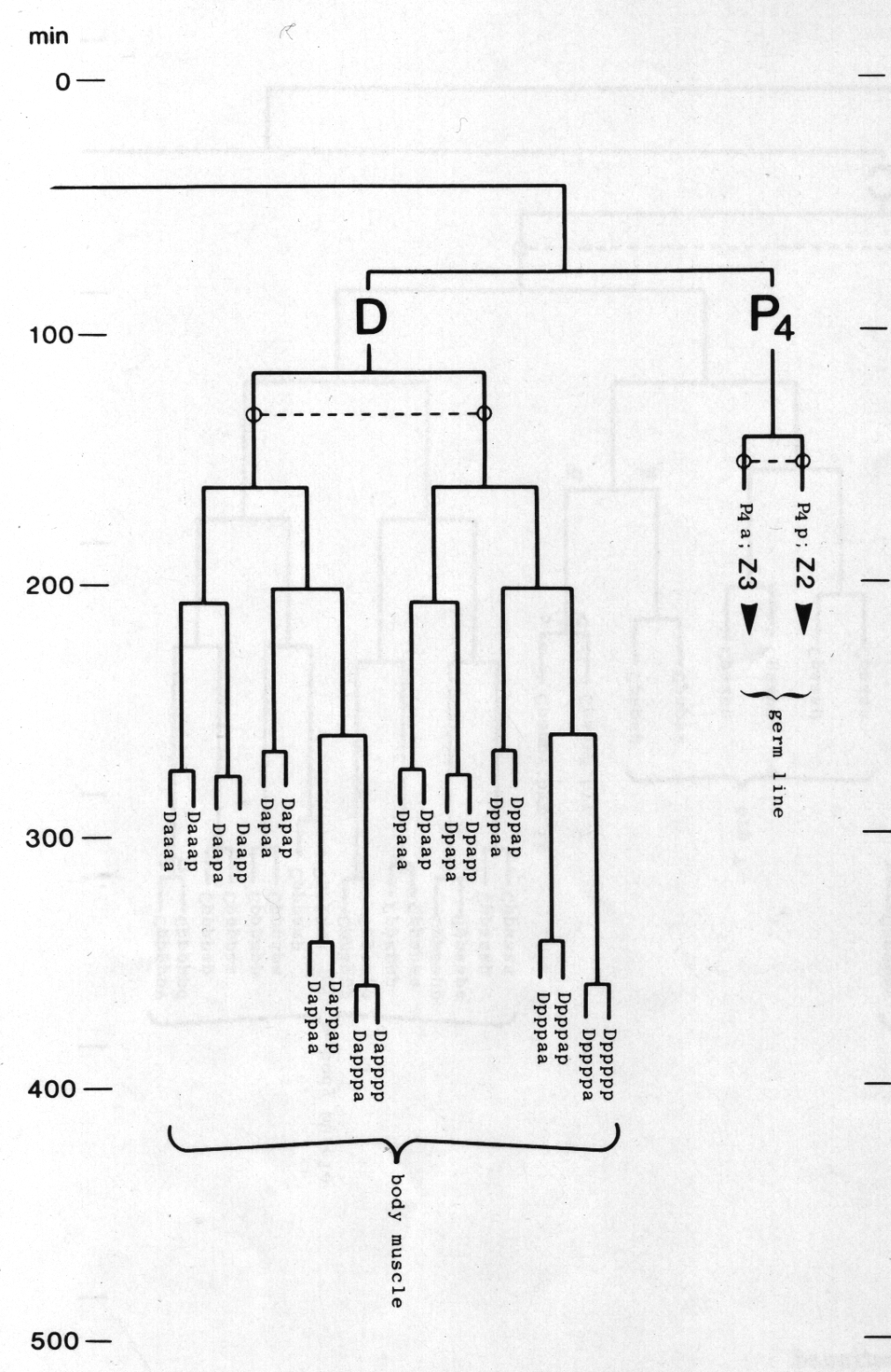
The development of the embryo has proved to be essentially invariant (Fig. 3- See complete Fig. 3 here). There seems to be no naturally occurring indeterminacy like that found postembryonically in the gonad (Kimble and Hirsh, 1979), the ventral hypodermis, and the male tail (Sulston and Horvitz, 1977).
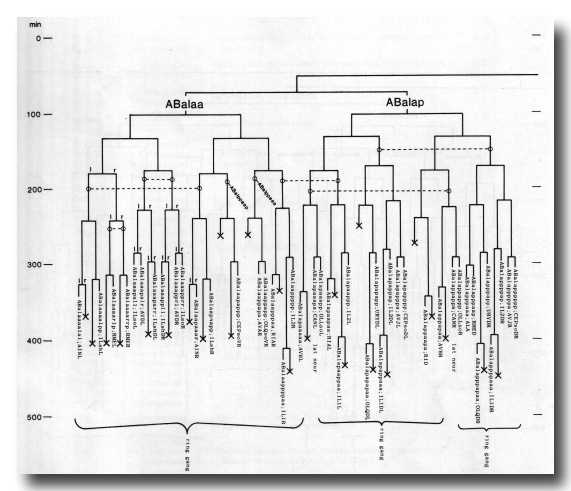
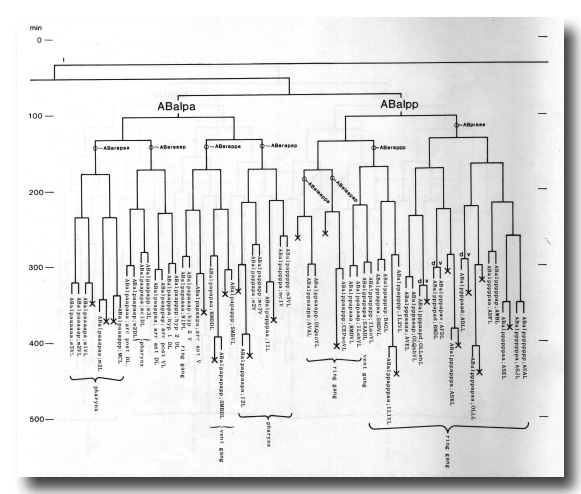
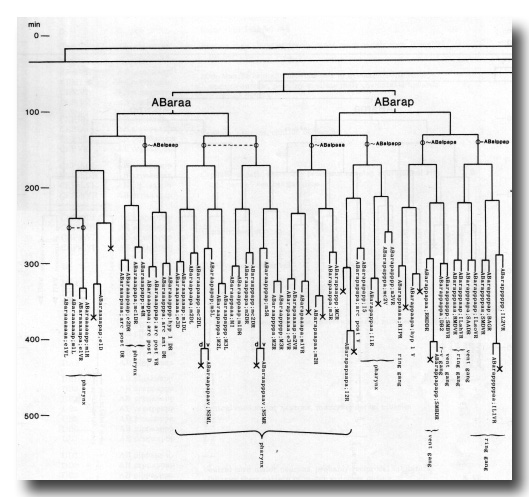
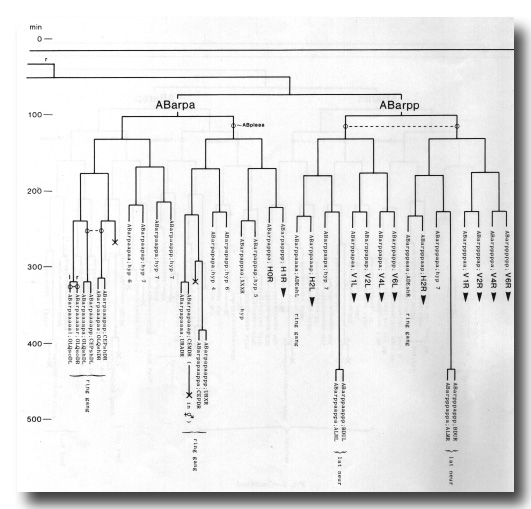
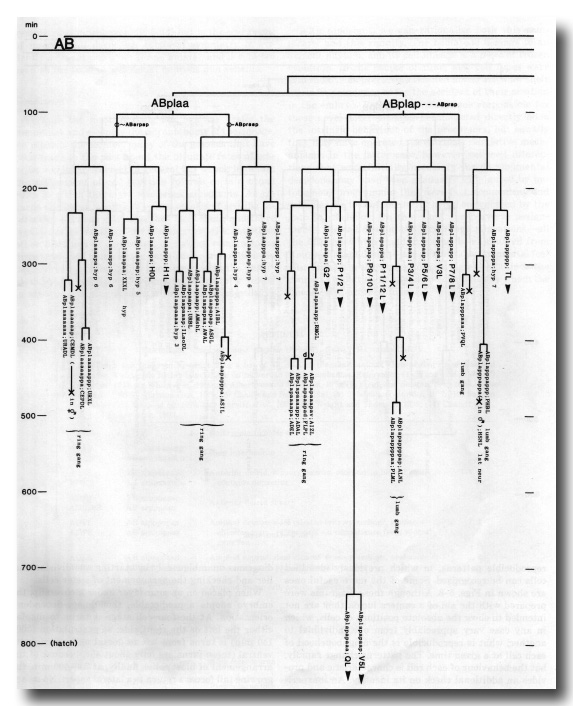
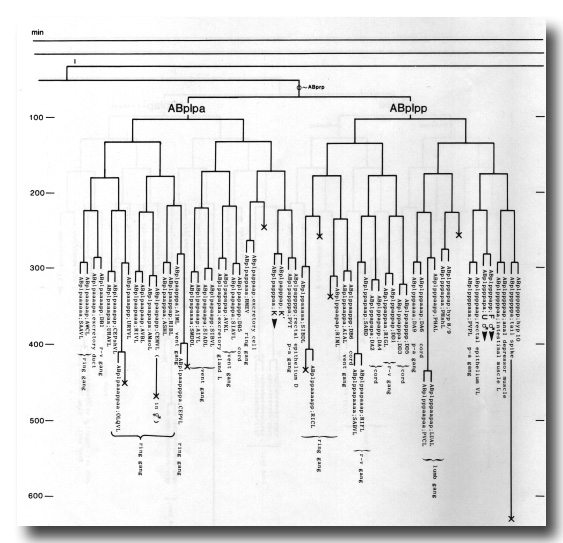
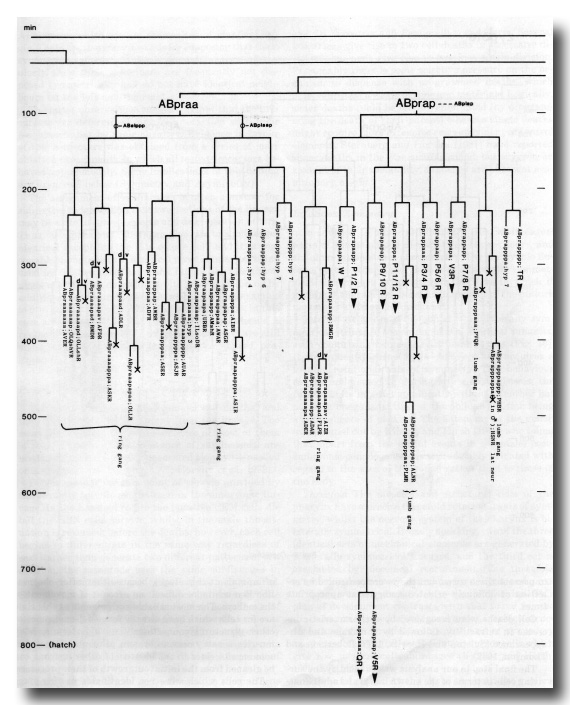

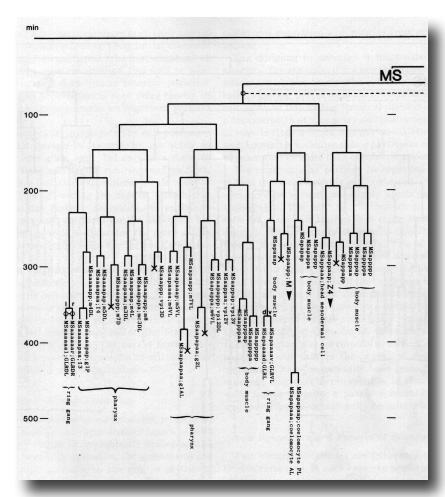
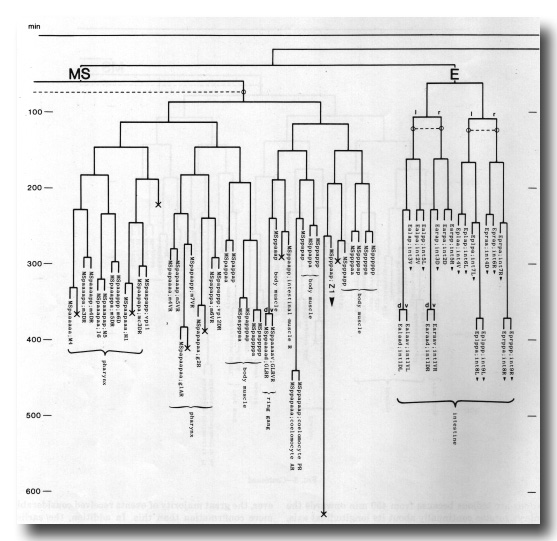
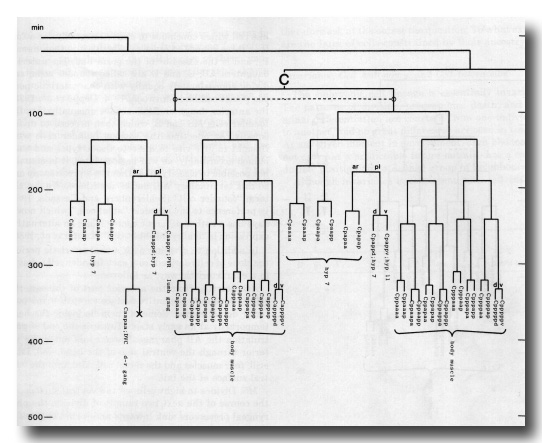
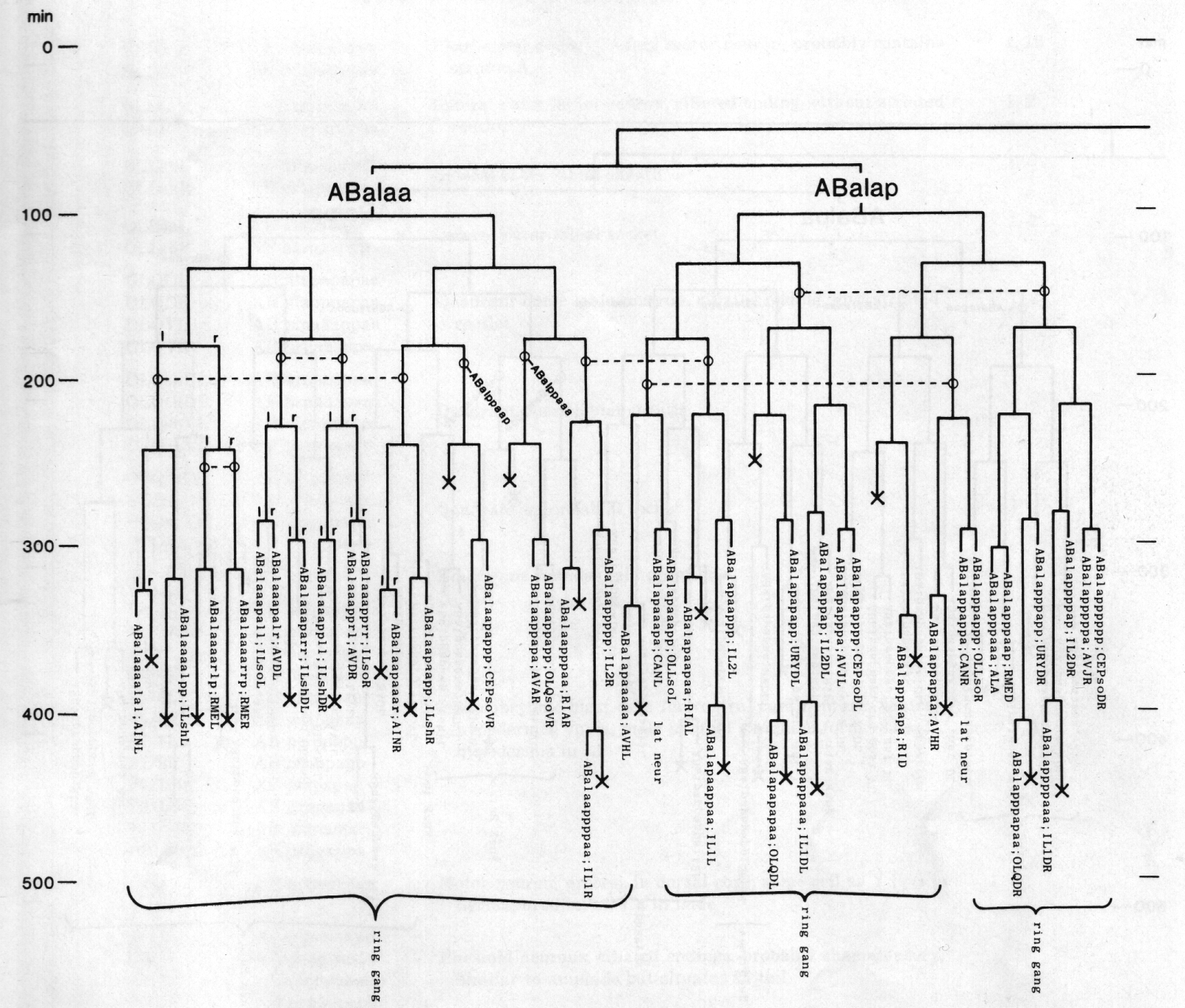
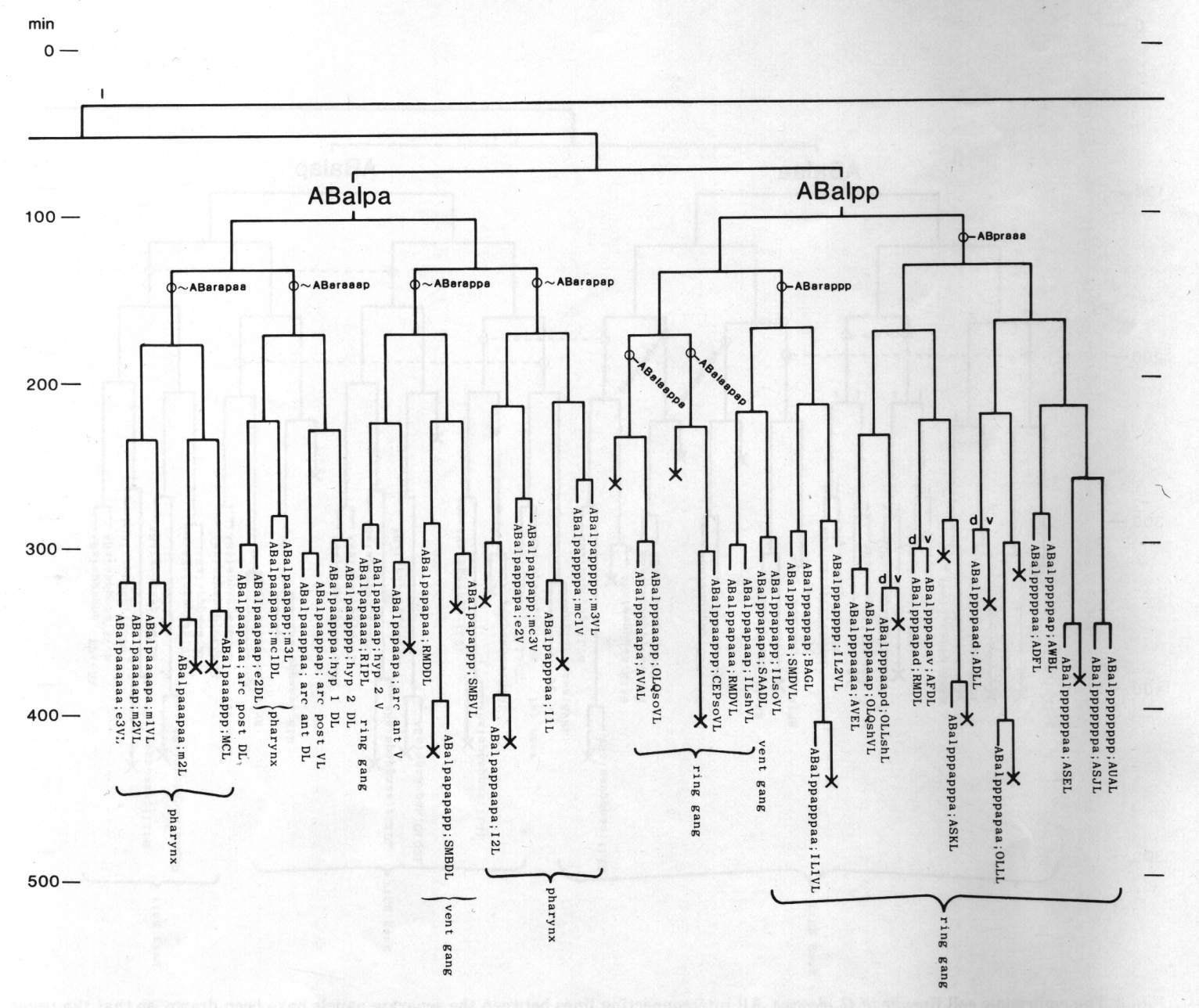
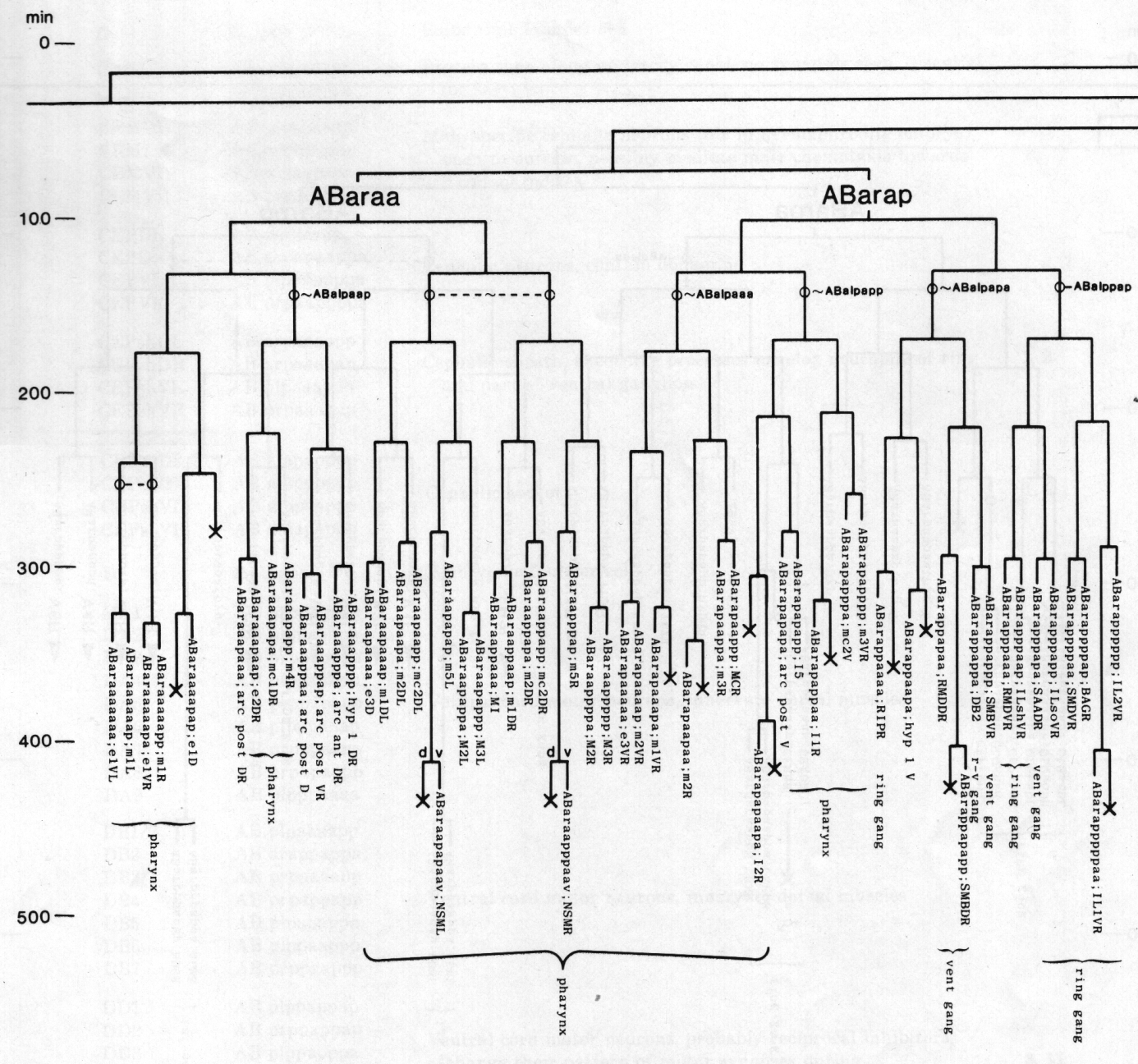
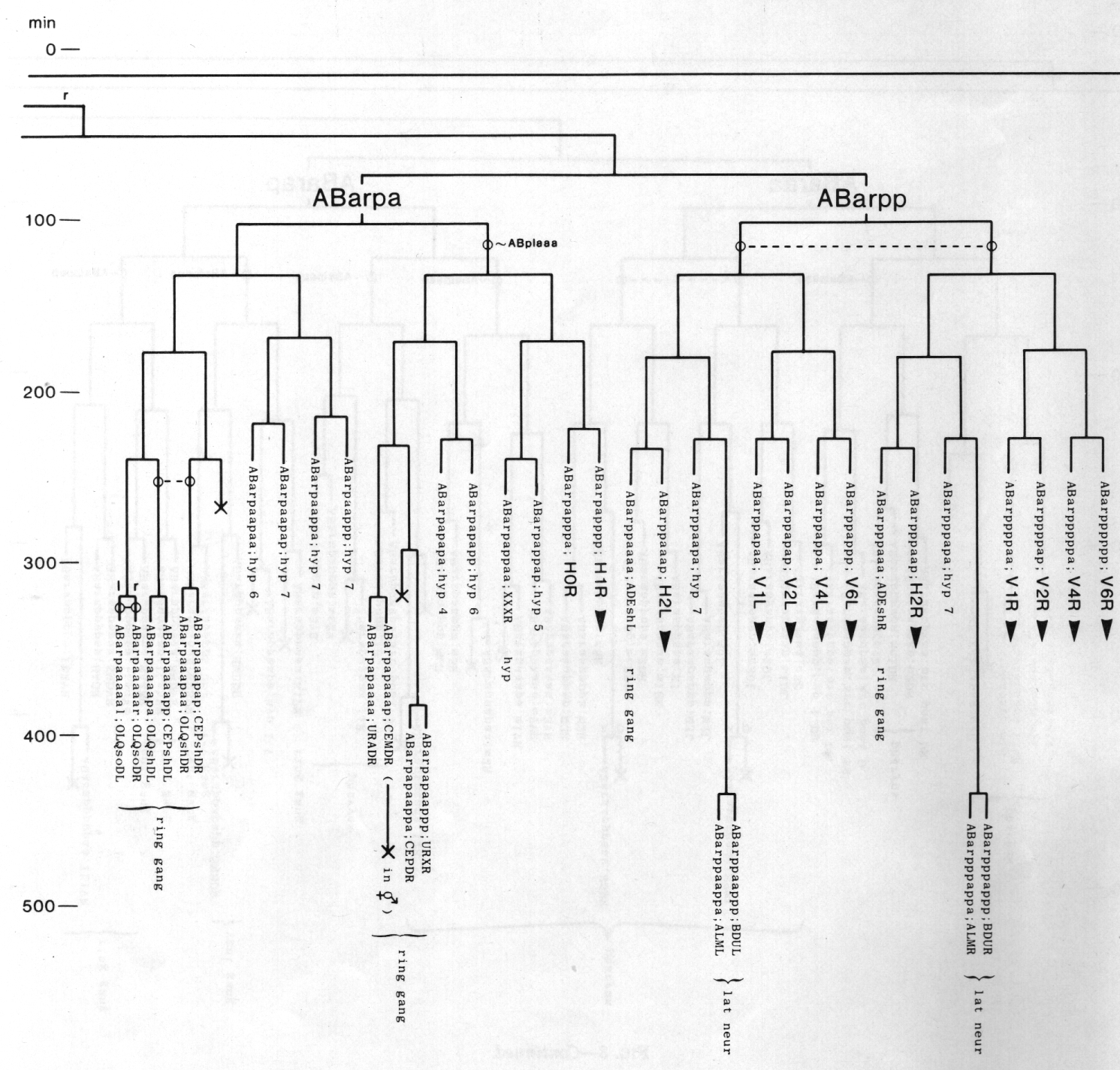
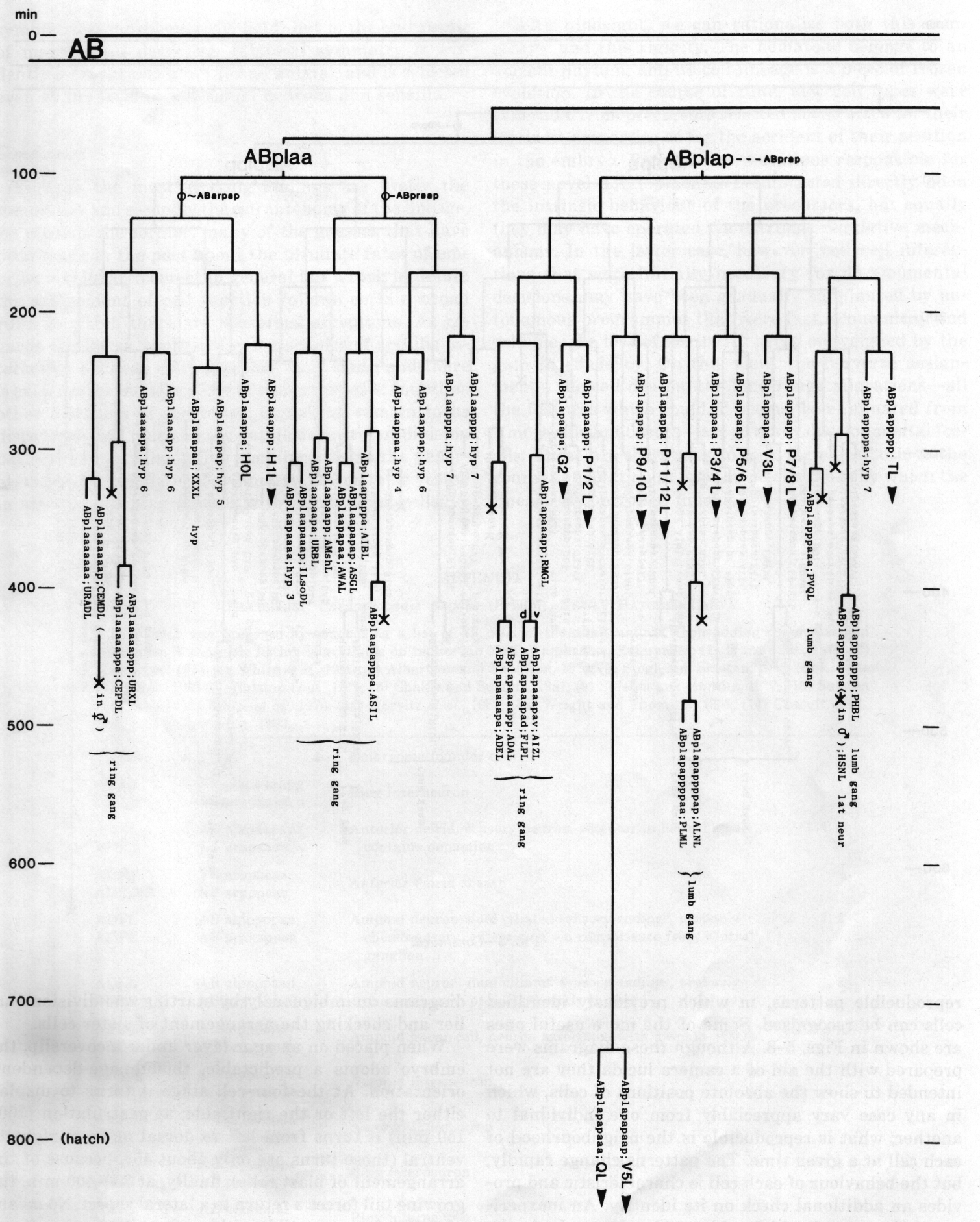
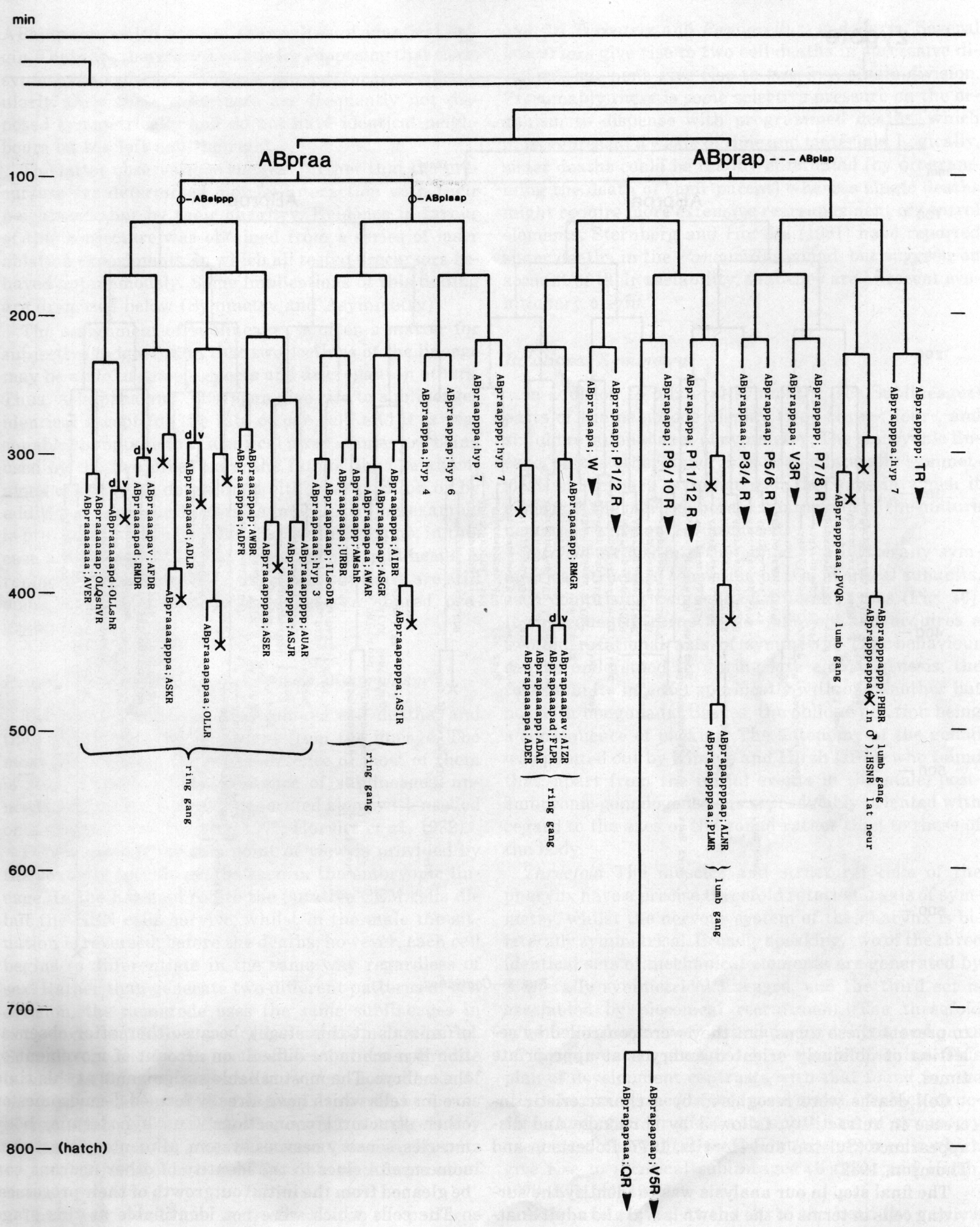
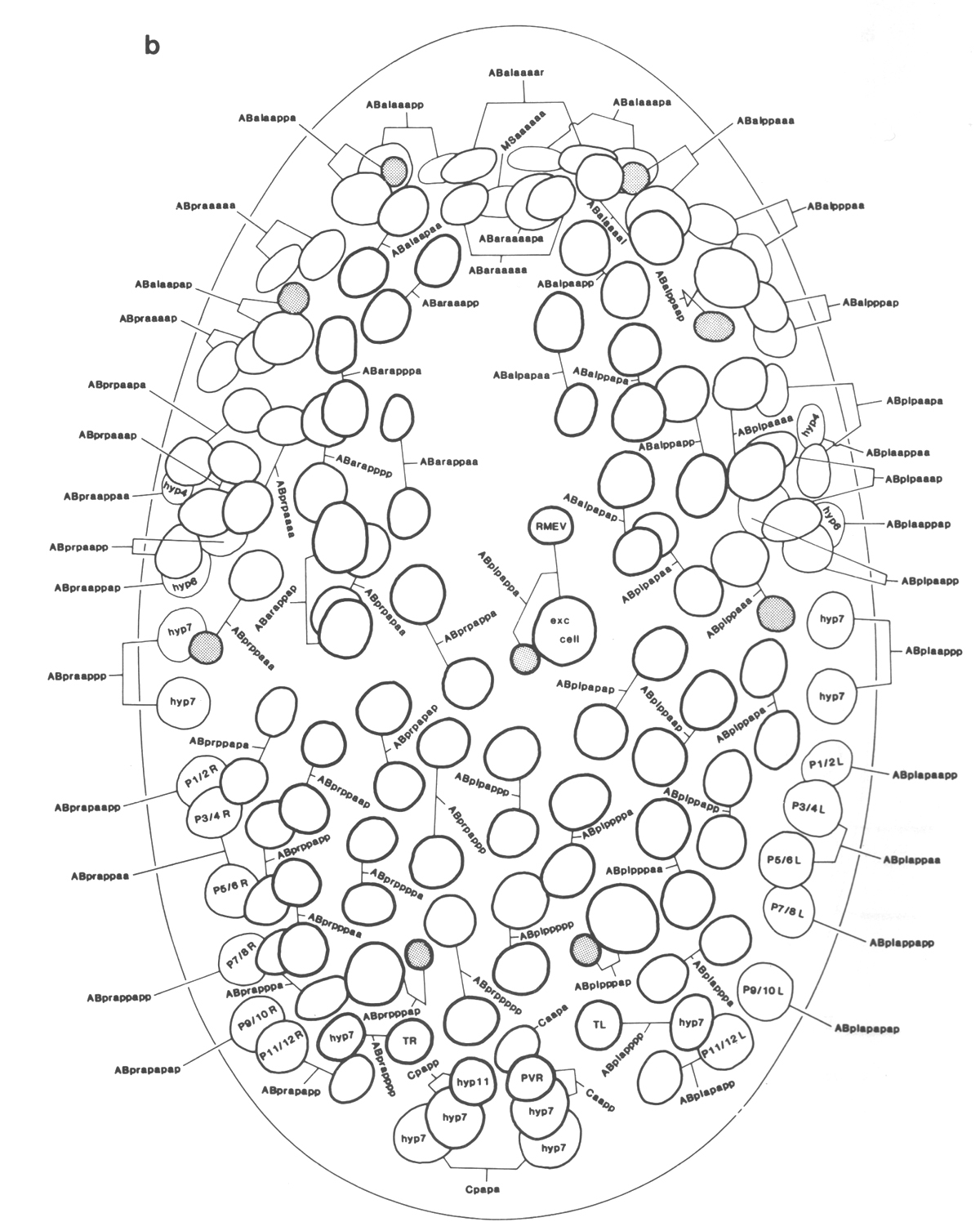
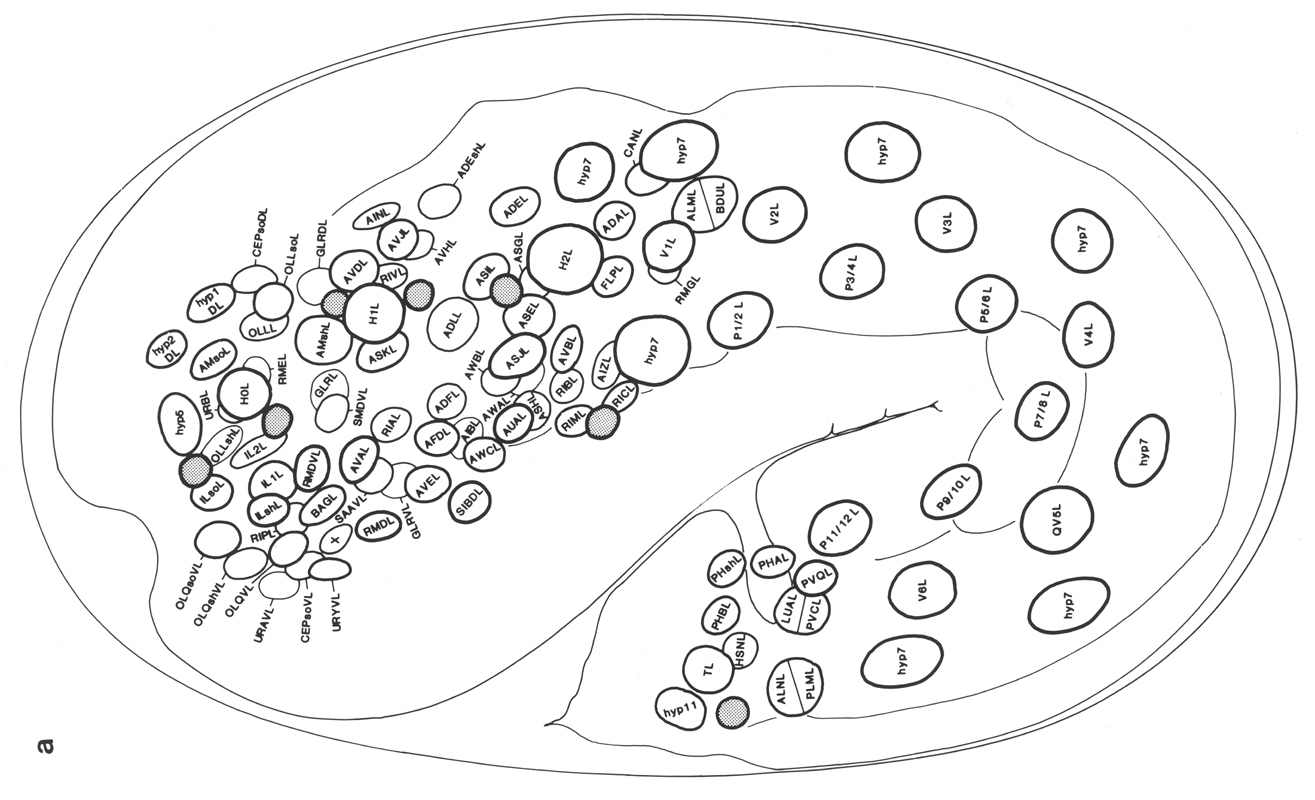
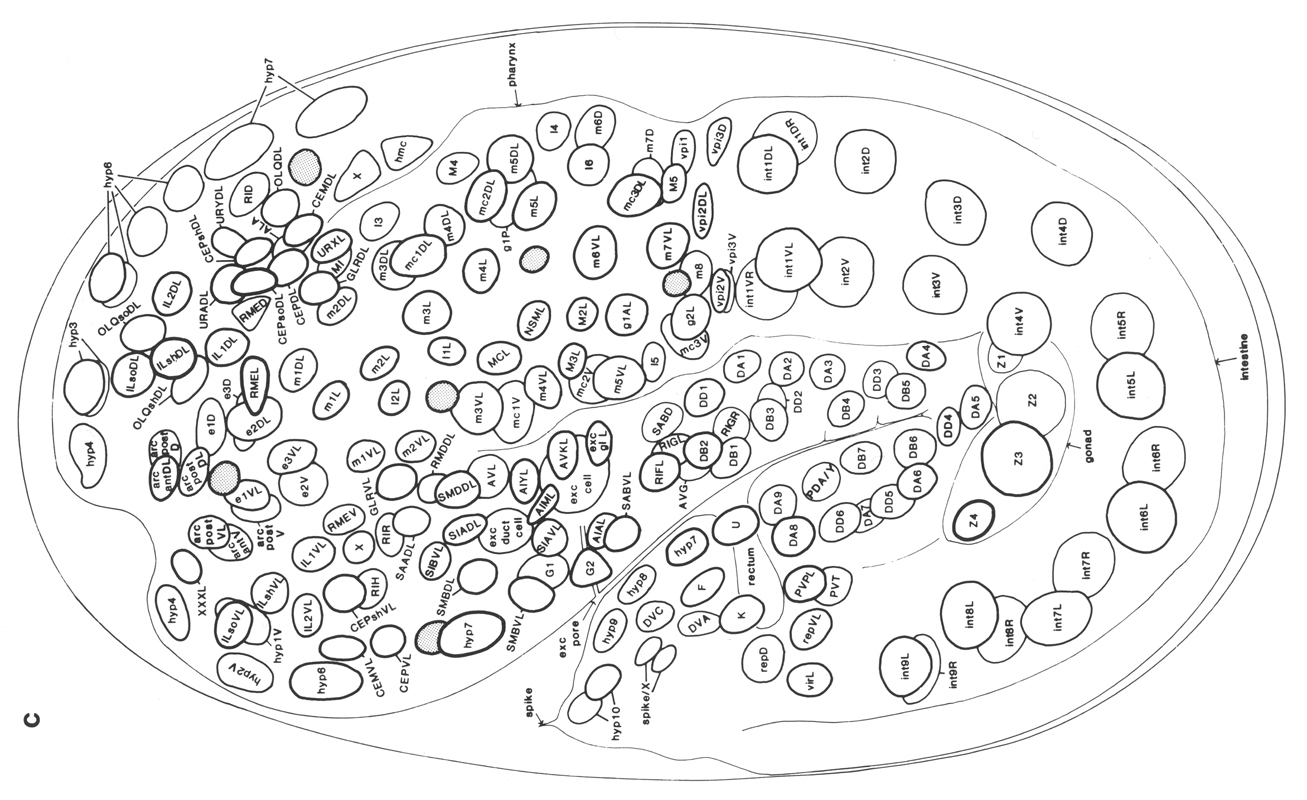
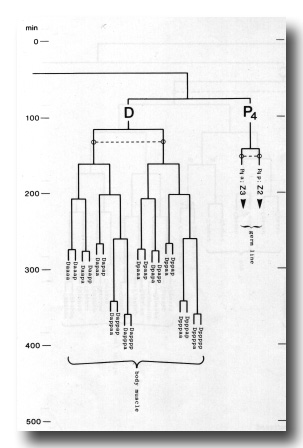 Figure 3. The embryonic cell linage of C. elegans. All interconnecting lines between the separate panels have been drawn, so that the pages can be copied, trimmed, and pasted together to give a complete chart. Vertical axis represents time at 20°C, from 0 min at first cleavage to 800 min at hatching. Many of the observations were made on eggs which were developing at slightly different rates (due to temperature variation and the effect of prolonged illumination); these primary results were normalised, by means of certain prominent cell divisions, to the course of events in eggs which were kept at 20°C and viewed infrequently. The precise times of individual events were not our primary concern, and should not be taken too seriously; the likely error varies from ±10% at the beginning of the lineage to ±2% at 400 min. Horizontal axis represents the direction of cell division. The majority of divisions have a marked anterior-posterior bias, and are shown with anterior to the left and posterior to the right, without any label. Only when this would lead to ambiguity in naming the daughters is an alternative direction indicated (l, left; r, right; d, dorsal; v, ventral); thus, our system is taxonomic rather than fully descriptive. The natural variation seen suggests that the precise direction of cell divisions is unimportant, at least in later development. Note that the daughters of a left-right division are not necessarily bilaterally symmetrical: for example, all the cells derived from ABalaaapa lie on the left of the animal and the right-hand daughters lie nearer the midline. Each terminal branch of the embryonic lineage is labelled either with X (indicating cell death; the position of the X on the time axis indicates the time of maximum refractility) or with a lineage name followed by a functional name. Large arrowheads denote cells which divide postembryonically, and small arrowheads denote nuclei which divide postembryonically, Symbols O and --- link precursors which give rise to bilaterally symmetrical groups of cells, the symbol ~ being included for cases of imperfect symmetry (see: Sublineages; Symmetry and Asymmetry), cord, ventral cord; gang, ganglion; lumb, lumbar; d-r, dorsorectal; p-a, preanal, r-v, retrovesicular; lat neur, isolated neuron lying laterally (WA editor's note: MSpappal is shown as MSpappaa in this diagram and MSpappar is shown as MSpappap (cf. Fig 15)(R. Lee pers. comm., and Richards et al., 2013.)
Figure 3. The embryonic cell linage of C. elegans. All interconnecting lines between the separate panels have been drawn, so that the pages can be copied, trimmed, and pasted together to give a complete chart. Vertical axis represents time at 20°C, from 0 min at first cleavage to 800 min at hatching. Many of the observations were made on eggs which were developing at slightly different rates (due to temperature variation and the effect of prolonged illumination); these primary results were normalised, by means of certain prominent cell divisions, to the course of events in eggs which were kept at 20°C and viewed infrequently. The precise times of individual events were not our primary concern, and should not be taken too seriously; the likely error varies from ±10% at the beginning of the lineage to ±2% at 400 min. Horizontal axis represents the direction of cell division. The majority of divisions have a marked anterior-posterior bias, and are shown with anterior to the left and posterior to the right, without any label. Only when this would lead to ambiguity in naming the daughters is an alternative direction indicated (l, left; r, right; d, dorsal; v, ventral); thus, our system is taxonomic rather than fully descriptive. The natural variation seen suggests that the precise direction of cell divisions is unimportant, at least in later development. Note that the daughters of a left-right division are not necessarily bilaterally symmetrical: for example, all the cells derived from ABalaaapa lie on the left of the animal and the right-hand daughters lie nearer the midline. Each terminal branch of the embryonic lineage is labelled either with X (indicating cell death; the position of the X on the time axis indicates the time of maximum refractility) or with a lineage name followed by a functional name. Large arrowheads denote cells which divide postembryonically, and small arrowheads denote nuclei which divide postembryonically, Symbols O and --- link precursors which give rise to bilaterally symmetrical groups of cells, the symbol ~ being included for cases of imperfect symmetry (see: Sublineages; Symmetry and Asymmetry), cord, ventral cord; gang, ganglion; lumb, lumbar; d-r, dorsorectal; p-a, preanal, r-v, retrovesicular; lat neur, isolated neuron lying laterally (WA editor's note: MSpappal is shown as MSpappaa in this diagram and MSpappar is shown as MSpappap (cf. Fig 15)(R. Lee pers. comm., and Richards et al., 2013.)
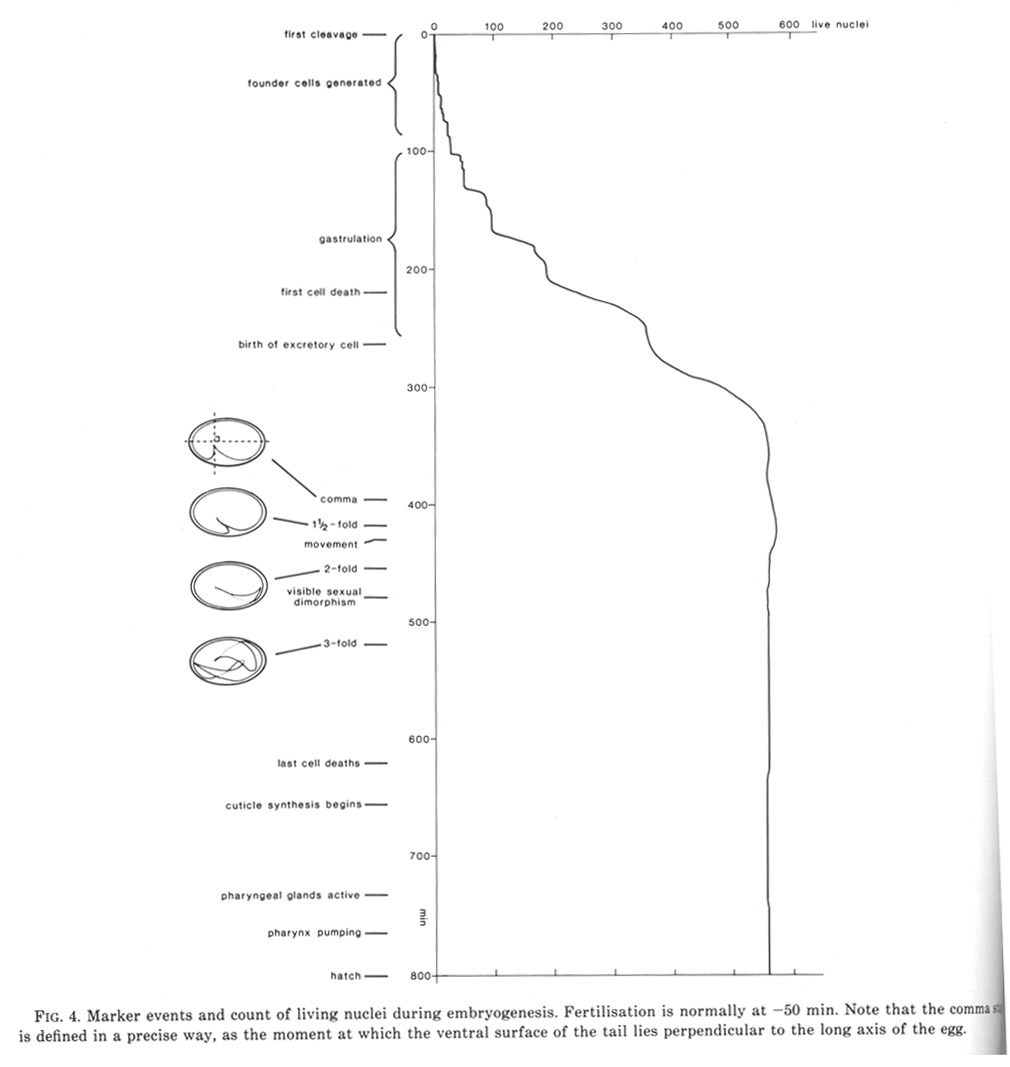
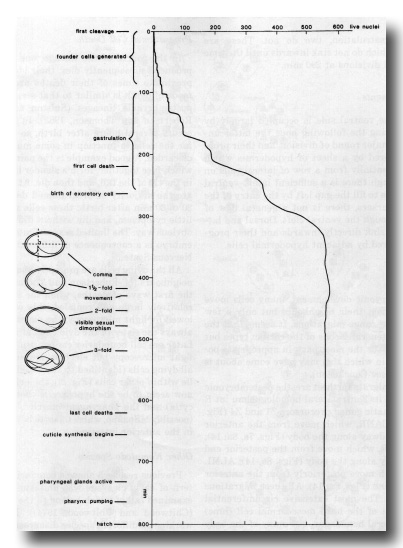 Figure 4. Marker events and count of living nuclei during embryogenesis. Fertilisation is normally at -50 min. Note that the comma stage is defined in a precise way, as the moment at which the ventral surface of the tail lies perpendicular to the long axis of the egg.
Figure 4. Marker events and count of living nuclei during embryogenesis. Fertilisation is normally at -50 min. Note that the comma stage is defined in a precise way, as the moment at which the ventral surface of the tail lies perpendicular to the long axis of the egg.
Cell Divisions and Cell Movement
The majority of cell divisions take place during the first half of embryogenesis. During the second half, the embryo changes greatly in external appearance: it elongates more than threefold, moves actively, synthesises cuticle, and initiates pharyngeal pumping before breaking out of the egg. This does not mean, however, that in the first half of embryogenesis there is no differentiation. On the contrary, by 430 min (the stage shown in figure 8), gastrulation and organogenesis are complete and the majority of the desmosomes seen in the L1 larva have already been made; subsequent events involve principally stretching and functional maturation of cells.
The Founder Cells
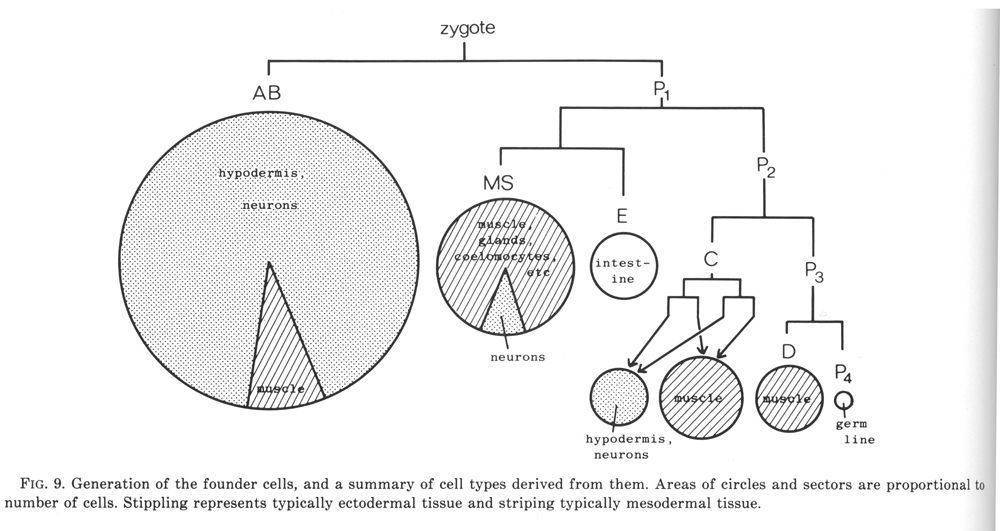
The fertilised egg cleaves into a larger anterior and a smaller posterior daughter. The latter (P1) is a stem- like cell which continues to cleave unequally for a further three rounds of division (figure 9); it is then named P4, and is the ancestor of the germ line. The anterior daughters AB, C, and D are called founder cells, and divide approximately equally with characteristic periods (Chitwood and Chitwood, 1974; Deppe at al, 1978); the anterior daughter EMS divides unequally into the founder cells MS and E, which then proceed to divide equally. At one time the various founder cells were thought to give rise to discrete tissue types and were named accordingly; as will be seen later, it is actually not possible to classify all of them in a precise way and so this terminology will not be used here. We use the term "founder cell" (Schierenberg and Cassada, 1982) in preference to the classical "stem cell," which nowadays has a rather different connotation; an alternative expression is "embryonic blast cell" (Laufer et al, 1980).
In addition to dividing with a characteristic period, the clone of cells derived from each founder cell behaves in a characteristic way, as follows:
AB. Starting from the anterior part of the egg, the cells spread over the entire surface except for the posterior dorsal region. Some of them lie inside the head temporarily at an early stage. Towards the end of gastrulation the AB pharyngeal precursors enter the interior through the ventral side of the head, and, later still, four muscles and the rectal cells sink into the ventral surface of the tail.
MS. Divides to eight cells on the ventral surface. In the course of the next two rounds of division the pharyngeal precursors sink inwards and form two rows in the head; meanwhile, the body muscle, coelomocyte, and somatic gonad precursors insinuate themselves between the intestine and the surface layer of AB cells.
E. Generates only intestine. Divides ventrally into two cells, which are the first to enter the interior in the course of gastrulation. Further division leads to a cylinder of cells with distinctively granular cytoplasm lying along the body axis.
C. Starting from the posterior dorsal side of the embryo, the cells spread anteriorly and posteriorly. The most posterior ones are body muscle precursors, which travel round to the ventral side and enter the interior immediately after D. Most of the remaining cells are employed in forming the dorsal hypodermis over the posterior two-thirds of the body; their nuclei migrate contralaterally (see Migrations).
D. Generates only body muscle. Enters the interior after division to four cells, at the same time as MS(a/p)p.
P4. Generates only germ line. Enters the interior after E, and divides into two cells which extend lobes into the intestine (figure 17).
Gastrulation
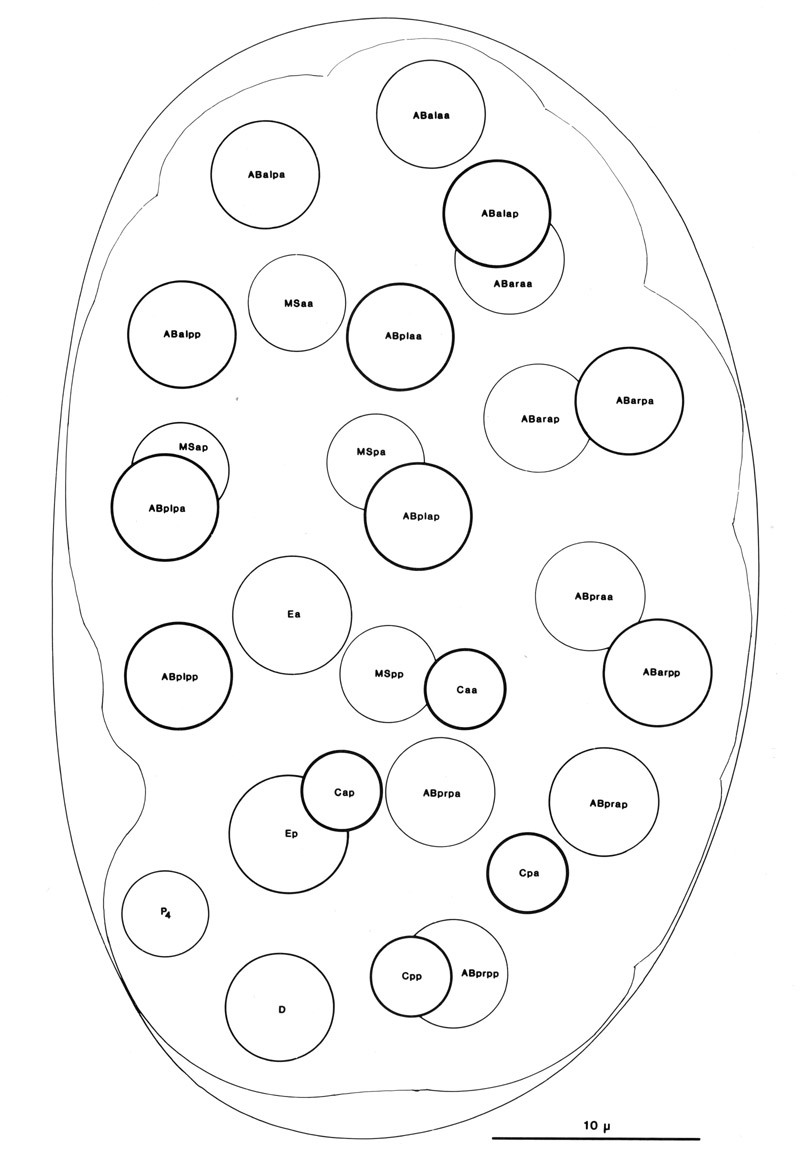
At 100 min after first cleavage, when the egg comprises 28 cells, gastrulation begins (figure 5). The first cells to enter the interior are Ea and Ep, which constitute the endoderm; they sink inwards from the ventral side, near the posterior end of the embryo. Next, at 120-200 min, are P4 and the progeny of MS. The entry zone widens and lengthens, spreading first posteriorly as most of the remaining myoblasts (derived from C and D) enter (180-230 min), and then anteriorly as the AB-derived part of the pharynx enters (210-250 min). The ventral cleft closes from posterior (230 min) to anterior (290 min). As gastrulation proceeds, the clone of E cells and the precursors of the pharynx form a central cylinder, while the body myoblasts insinuate themselves between this cylinder and the outer layer of cells. Although most of the myoblasts enter the body cavity during gastrulation, two do not. These are ABp(l/r)pppppa, which do not sink inwards until the time of their terminal divisions at 290 min.
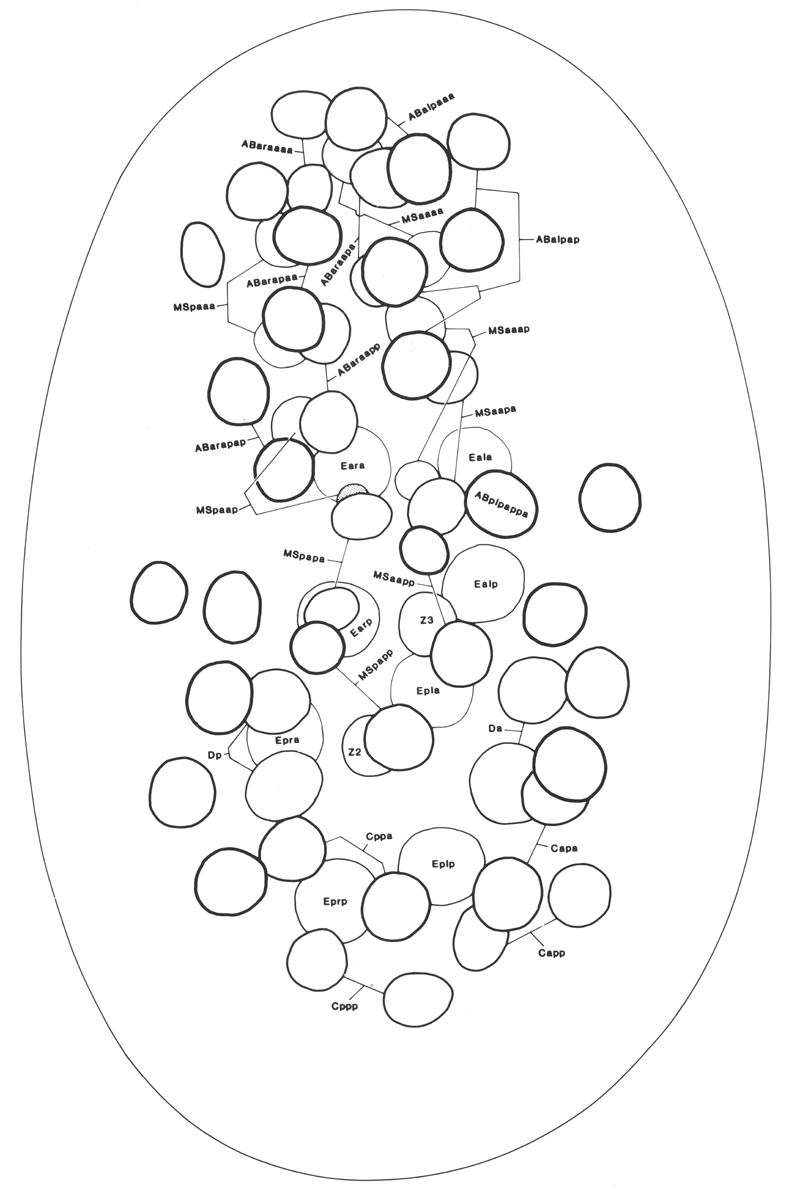
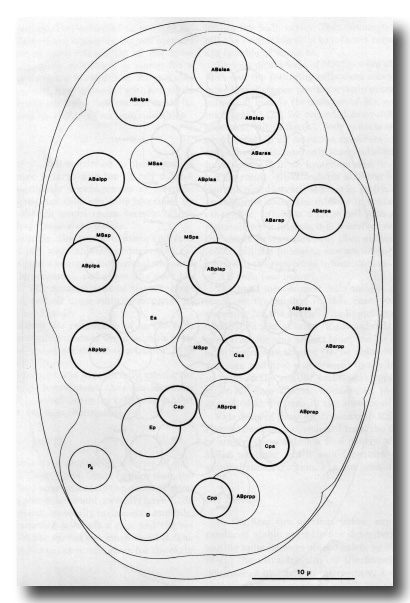 Figures 5-8. Drawings of embryos. Circles and ovals represent nuclei, traced by means of a camera lucida, the thickness of the lines being inversely related to depth; outlines of the egg, embryo, and internal structures are traced with thin lines (regardless of depth). Anterior is towards the top of the page. Dying nuclei are stippled.
Figures 5-8. Drawings of embryos. Circles and ovals represent nuclei, traced by means of a camera lucida, the thickness of the lines being inversely related to depth; outlines of the egg, embryo, and internal structures are traced with thin lines (regardless of depth). Anterior is towards the top of the page. Dying nuclei are stippled.
Figure 5. Embryo, 100 min, left dorsal aspect; all nuclei included; cf. Fig. 2b. This stage has already been well characterised (Krieg et al, 1978), and the observer quickly learns to recognise all the nuclei; it is a useful starting point both for lineages and for ablation experiments. An embryo in the orientation shown will present a dorsal aspect until it turns at 350 min; an embryo with the MS cells uppermost will present a ventral aspect. The intestinal precursors are entering the interior, leaving a characteristic depression on the ventral side.
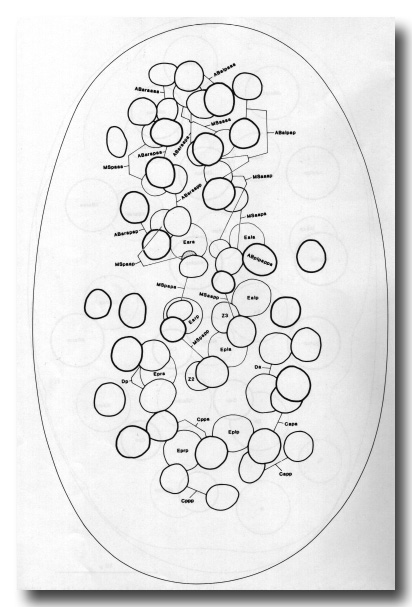 Figure 6. Embryo, 200 min, ventral aspect, late gastrulation. The orientation of the egg can be recognised by the prominent ventral cleft; cf. Fig. 2c. All progeny of MS, E, D, and P4 are shown, together with AB-derived pharyngeal precursors and C-derived myoblasts; other AB progeny (unlabelled) are included for perspective. MSaa and MSpa have each formed a chain of eight cells, of which all but the posterior two have entered the interior. The derivatives of ABaraap have also entered, and the remainder of the AB pharyngeal precursors will soon follow. Posteriorly, the mesodermal precursors derived from C and D are also entering the cleft. The germ line and the intestine can be seen lying more dorsally. The division of ABplpappa (earlier than its neighbours and very unequal) is a useful time point. Embryos of this age, but viewed from the dorsal side, were used as starting points for lineages of the dorsal pharynx and dorsal body muscle.
Figure 6. Embryo, 200 min, ventral aspect, late gastrulation. The orientation of the egg can be recognised by the prominent ventral cleft; cf. Fig. 2c. All progeny of MS, E, D, and P4 are shown, together with AB-derived pharyngeal precursors and C-derived myoblasts; other AB progeny (unlabelled) are included for perspective. MSaa and MSpa have each formed a chain of eight cells, of which all but the posterior two have entered the interior. The derivatives of ABaraap have also entered, and the remainder of the AB pharyngeal precursors will soon follow. Posteriorly, the mesodermal precursors derived from C and D are also entering the cleft. The germ line and the intestine can be seen lying more dorsally. The division of ABplpappa (earlier than its neighbours and very unequal) is a useful time point. Embryos of this age, but viewed from the dorsal side, were used as starting points for lineages of the dorsal pharynx and dorsal body muscle.
Later Cell Movements
At 250 min the ventral side is occupied largely by neuroblasts. During the following hour the latter undergo their last major round of division, and their progeny become covered by a sheet of hypodermis which grows circumferentially from a row of lateral cells on each side. Although there is a sufficient latero-ventral movement of cells to fill the gap left by the entry of the pharyngeal precursors, there is not a general flow of neural tissue through the ventral cleft. Dorsal and lateral neuroblasts sink directly inwards and their progeny become covered by adjacent hypodermal cells.
Migrations
As in postembryonic development, many cells move short distances past their neighbours but only a few embark upon long-range migrations. It seems that the lineage not only generates cells of the correct types but also places them, for the most part, in appropriate positions. The way in which this may have come about is suggested later (see Conclusion).
The cells that migrate furthest are: the postembryonic mesoblast M and its contralateral homologue mu int R (figure 16); the somatic gonad precursors Z1 and Z4 (figure 16); CANL and CANR, which move from the anterior end to a point midway along the body (Figs. 7a, 8a, 14); HSNL and HSNR, which move from the posterior end to a point midway along the body (Figs. 8a, 14); ALML and ALMR, which move posteriorly from the anterior end of the intestine (Figs. 8a, 14). All these migrations are longitudinal. The most extensive circumferential migration is that of the head mesodermal cell (hmc) and its contralateral homologue, but many of the body muscles also move circumferentially as they assemble into rows.
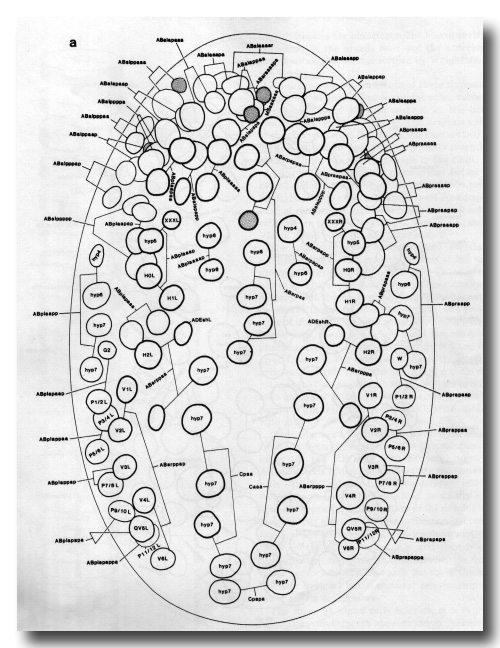 Figure 7a. Embryo, 260 min, dorsal aspect, superficial nuclei; cf. Fig. 2d. Landmarks: nuclei of hyp4-hyp7, cell deaths; time points: division of various neuroblasts. The dorsal hypodermal cells have very granular cytoplasm and form prominent transverse ridges. In figure 7 some licence has been allowed in depicting cell deaths, because of their importance in pattern recognition; in fact, they do not all become refractile simultaneously. WA editors' note: the positions of the descendants of Caaa and Cpaa are inadvertantly reversed in this fig (Loer C. et al, WBG 10(3)120).
Figure 7a. Embryo, 260 min, dorsal aspect, superficial nuclei; cf. Fig. 2d. Landmarks: nuclei of hyp4-hyp7, cell deaths; time points: division of various neuroblasts. The dorsal hypodermal cells have very granular cytoplasm and form prominent transverse ridges. In figure 7 some licence has been allowed in depicting cell deaths, because of their importance in pattern recognition; in fact, they do not all become refractile simultaneously. WA editors' note: the positions of the descendants of Caaa and Cpaa are inadvertantly reversed in this fig (Loer C. et al, WBG 10(3)120).
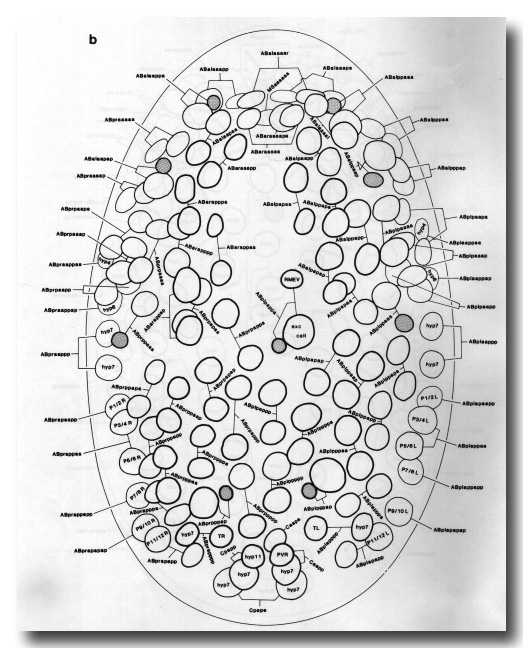 Figure 7b. Embryo, 270 min, ventral aspect, superficial nuclei; cf. Fig. 2e. Landmarks: excretory cell, cell deaths; time point: division of mother of excretory cell. The gap anterior to the excretory cell contains pharyngeal and buccal precursors which are entering the interior.
Figure 7b. Embryo, 270 min, ventral aspect, superficial nuclei; cf. Fig. 2e. Landmarks: excretory cell, cell deaths; time point: division of mother of excretory cell. The gap anterior to the excretory cell contains pharyngeal and buccal precursors which are entering the interior.
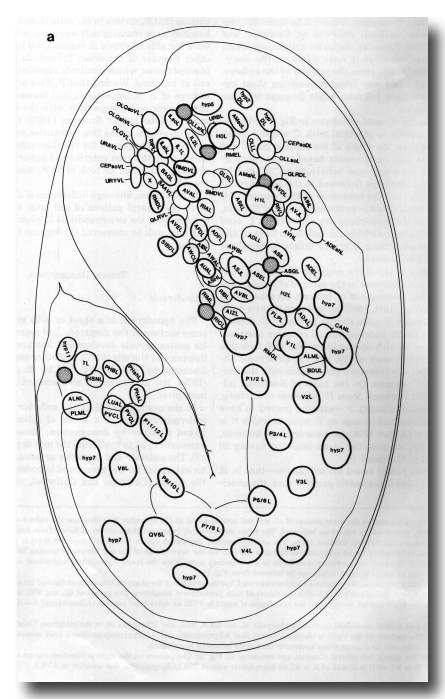
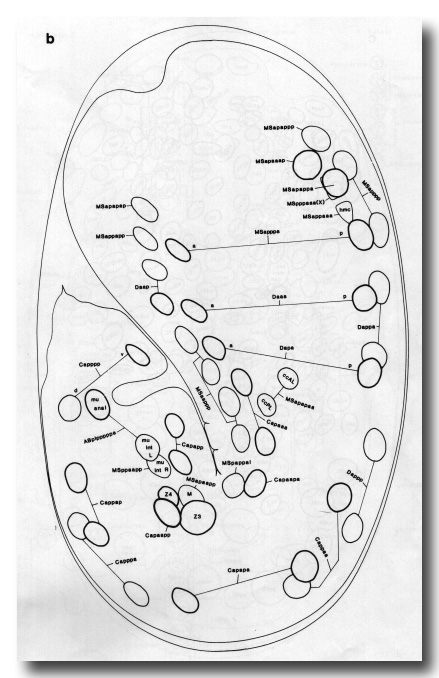
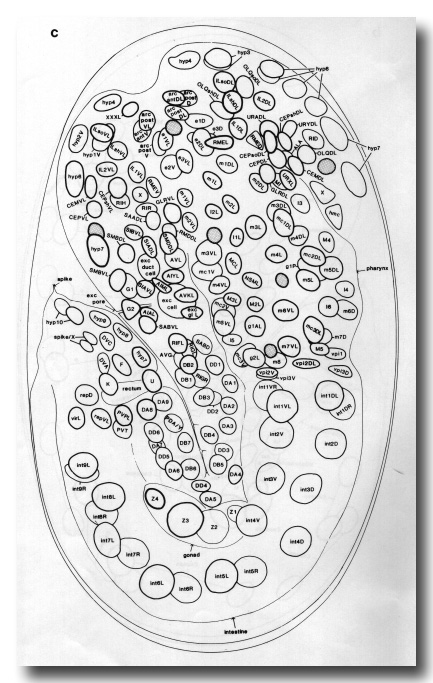 Figure 8. This figure illustrates the arrangement of all left and central nuclei at 430 min after first cleavage; the right-hand side is a mirror image of the left, except where otherwise indicated. The three parts roughly represent three planes of focus (from superficial to central), but there is considerable overlap between them. The anterior sensory depression (not a mouth opening -see text) is at the top and the lengthening tail, terminating in the spike, curves round to the left. On the ventral side of the tail the rectal opening has appeared, and on the ventral side of the head the excretory duct leads to the excretory pore. Since the major hypodermal migrations are complete at this stage, the ancestry of nuclei in hyp3-hyp11 can be inferred from figure 13.
Figure 8. This figure illustrates the arrangement of all left and central nuclei at 430 min after first cleavage; the right-hand side is a mirror image of the left, except where otherwise indicated. The three parts roughly represent three planes of focus (from superficial to central), but there is considerable overlap between them. The anterior sensory depression (not a mouth opening -see text) is at the top and the lengthening tail, terminating in the spike, curves round to the left. On the ventral side of the tail the rectal opening has appeared, and on the ventral side of the head the excretory duct leads to the excretory pore. Since the major hypodermal migrations are complete at this stage, the ancestry of nuclei in hyp3-hyp11 can be inferred from figure 13.
Figure 8a. Left lateral ectoderm. No midplane nuclei are included, but the outlines of the pharynx, intestine, and gonad are shown for reference. Nuclei which will soon divide are labelled with the names of both presumptive daughters. The parent of QL and V5L is named QV5L. The pattern on the right is identical, except that: the homologue of hyp11 is PVR; an extra hyp7 nucleus (ABarpaappp) lies dorsal to H2R; there is no hyp2 DR.
Figure 8b. Left and central mesoderm (excluding pharynx). M, mu int R, hmc, and MSpppaaa lie in the midplane. Unlabelled nuclei are in body muscles. The pattern on the right is identical, except that ABprpppppaa and ABprpppppap become a body muscle and the sphincter muscle, respectively, and lie slightly more anteriorly than their left-hand homologues.
Figure 8c. Intestine, gonad, left central pharynx, and ectoderm; cf. Fig. 2g. The pattern on the right is identical, except that: the homologue of G2 is W, of 16 is M1, of U is B, and of K is K'; an asymmetric neuron RIS (ABprpappapa) lies anterior to AVKR.
The group of cells which forms most of the dorsal hypodermis exhibits nuclear migration as opposed to cellular migration (see figure 10). These cells are born subdorsally, in two longitudinal rows; a cytoplasmic process grows circumferentially from each cell across the dorsal midline, and after a time the nucleus migrates along this process until it lies on the opposite side of the embryo. This type of migration is analogous to that seen postembryonically in the P cells (Sulston and Horvitz, 1977), and it is therefore interesting that mutations at two loci interfere with both processes (Sulston and Horvitz, 1981).
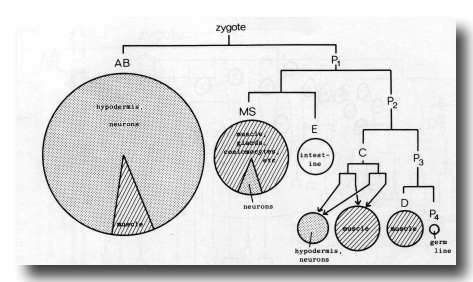 Figure 9. Generation of the founder cells, and a summary of cell types derived from them. Areas of circles and sectors are proportional to number of cells. Stippling represents typically ectodermal tissue and striping typically mesodermal tissue.
Figure 9. Generation of the founder cells, and a summary of cell types derived from them. Areas of circles and sectors are proportional to number of cells. Stippling represents typically ectodermal tissue and striping typically mesodermal tissue.
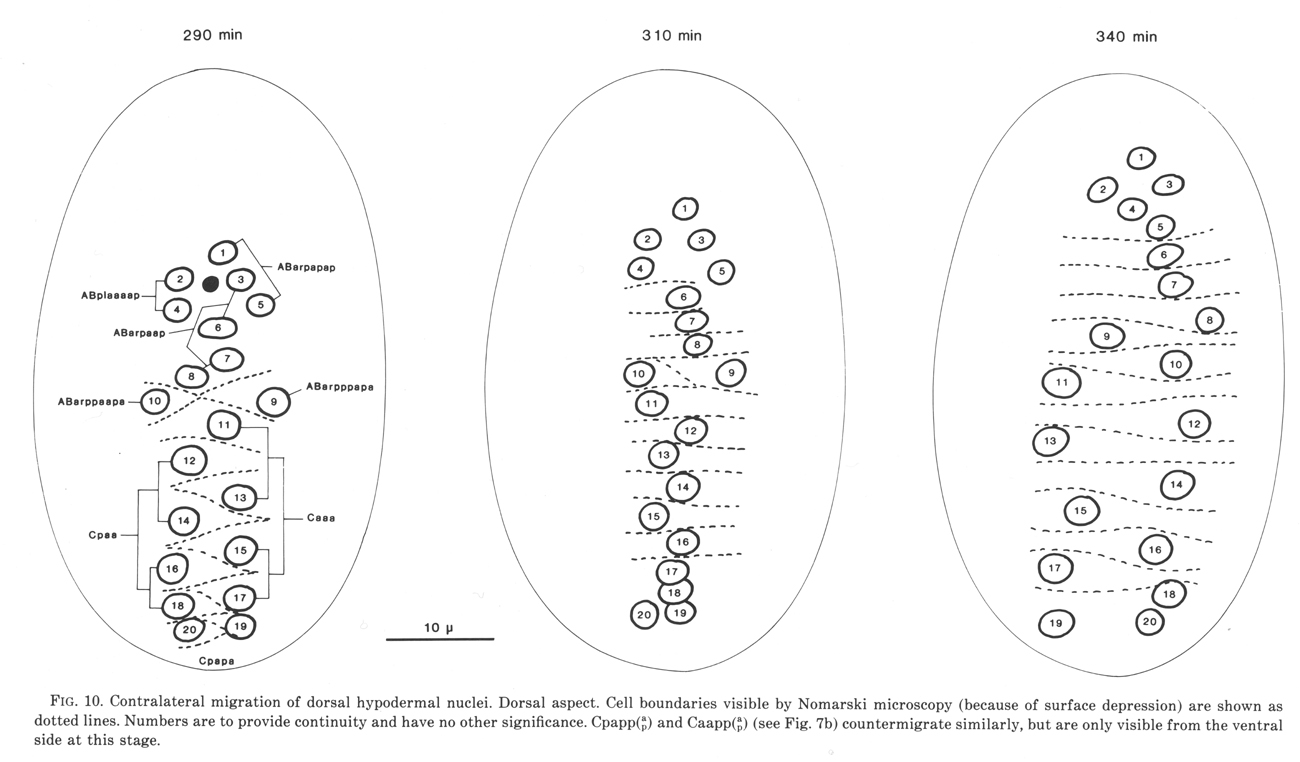
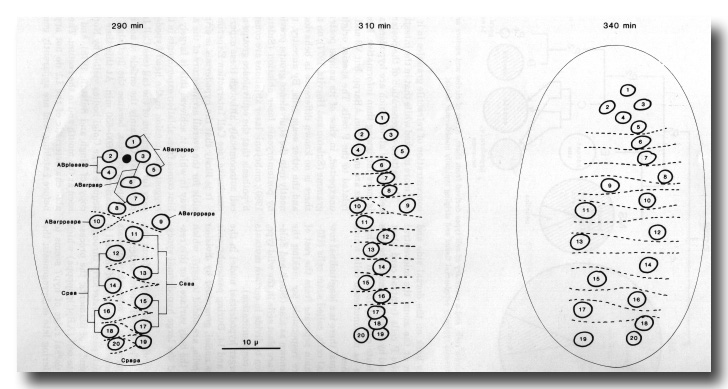 Figure 10. Contralateral migration of dorsal hypodermal nuclei. Dorsal aspect. Cell boundaries visible by Nomarski microscopy (because of surface depression) are shown as dotted lines. Numbers are to provide continuity and have no other significance. Cpapp(a/p) and Caapp(a/p) (see figure 7b) countermigrate similarly, but are only visible from the ventral side at this stage. WA editors' note: the positions of the descendants of Caaa and Cpaa are inadvertantly reversed in this fig (Loer C. et al, WBG 10(3)120).
Figure 10. Contralateral migration of dorsal hypodermal nuclei. Dorsal aspect. Cell boundaries visible by Nomarski microscopy (because of surface depression) are shown as dotted lines. Numbers are to provide continuity and have no other significance. Cpapp(a/p) and Caapp(a/p) (see figure 7b) countermigrate similarly, but are only visible from the ventral side at this stage. WA editors' note: the positions of the descendants of Caaa and Cpaa are inadvertantly reversed in this fig (Loer C. et al, WBG 10(3)120).
Programmed Cell Death
In the course of the lineage, one in six of all cells produced subsequently dies; their identity and the approximate times of their deaths are predictable. The mode of death is similar to that seen previously in the postembryonic lineages (Sulston and Horvitz, 1977; Robertson and Thomson, 1982). In some cases death occurs several hours after birth, so that it is possible for the cells to function in some manner before being discarded. A good example is the pair of tail spike cells, which fuse together, form a slender bundle of filaments in the tip of the tail, and then die. At the other extreme are the majority of programmed deaths, which occur 20 to 30 min after birth; these cells are born with very little cytoplasm, and die without differentiating in any obvious way. The limited sexual dimorphism seen in the embryo is a consequence of differential cell death (see Nervous System).
All the dying cells are promptly phagocytosed by their neighbours (cf. Robertson and Thomson, 1982). During the first wave of deaths, when the surviving cells are relatively large and rounded, this process can be followed by light microscopy; the engulfing cell is almost always the sister of the dying cell at this time (Fig. 2e). Later on, cell boundaries cannot usually be resolved by light microscopy but electron micrographs show that all dying cells (identified by their high electron density) lie within other cells (Fig. 2i); the principal phagocytes now seem to be the hypodermis (both seams and syncytia) and the pharyngeal muscles, though one death (possibly MSpaapp, which tends to be delayed) was found in the anterior intestine.
Other Nematode Species
Previous research showed convincingly that the pattern of early cleavage was uniform in the nematodes examined (all of which belong to the class Secernentea (Chitwood and Chitwood, 1974). However, authors working on different species disagreed about the tissues to which certain founder cells gave rise; these disagreements may have been due either to the difficulties of interpreting observations on fixed specimens or to genuine differences between the species.
The most interesting discrepancy is in the origin of the somatic gonad, which was reported to arise from the founder cell P4 in the nematodes Turbatrix aceti (Pai, 1927), Ascaris megalocephala (Boveri, 1892; but not sustained in later reports), and Bradynema rigidum (zur Strassen, 1959). Given the consistency of the early cleavage pattern of nematode embryos, a switch in the origin of such a vital tissue would be surprising indeed. Other discrepancies are in the fates of C progeny (said to be only five, and exclusively ectodermal, in T. aceti (Pai, 1927)) and D progeny (reported to form the rectum, by several authors; reviewed by Chitwood and Chitwood (1974)). However, the latter assignment seems to be hypothetical, since it rests solely on the observation that the D cells enter the interior of the embryo. In order to go some way towards resolving these uncertainties, we have followed a few lineages in other nematode species.
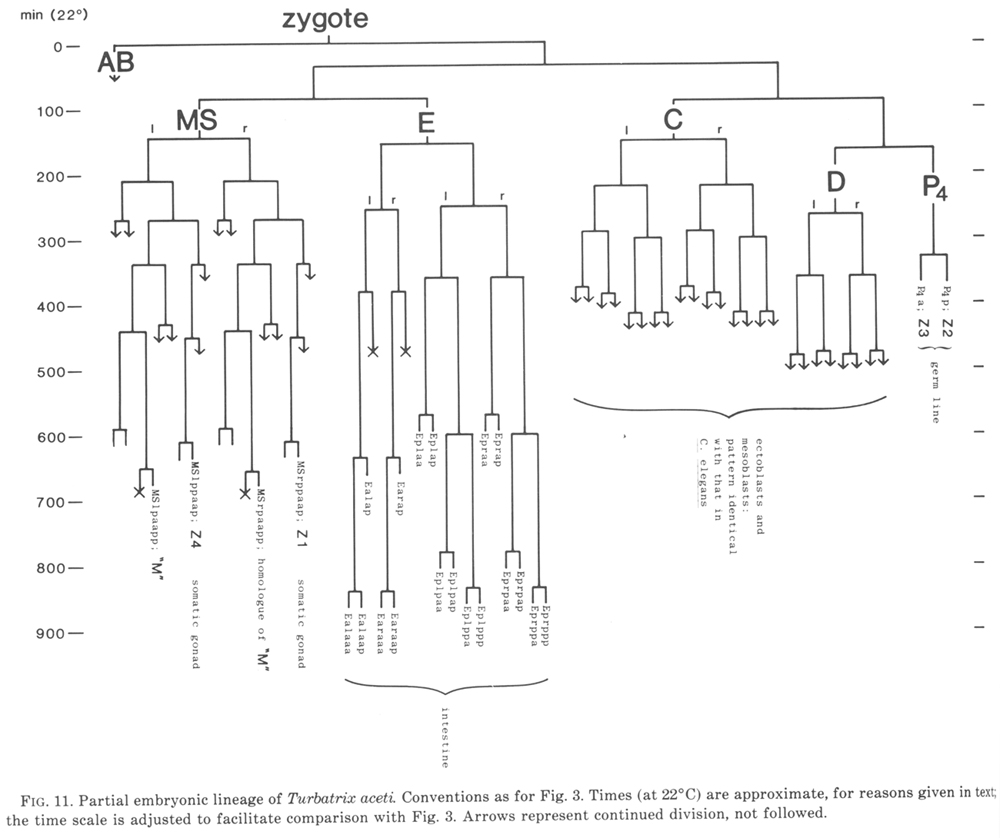
The results for T. aceti are shown in fig. 11. We find that this nematode is identical with C. elegans in the following respects: the origin of the somatic gonad and the germ line (cell assignments based on morphology at hatching); the origin and behaviour of "M" and its contralateral homologue (followed to the equivalent of 320 min, cf. figure 16); the behaviour of the progeny of C and D (followed explicitly to the equivalent of 230 min, by which time the mesoblasts are distinctive). T. aceti differs from C. elegans within the endoderm lineage, and it will not be surprising if it also differs in details of the MS and AB lineages.
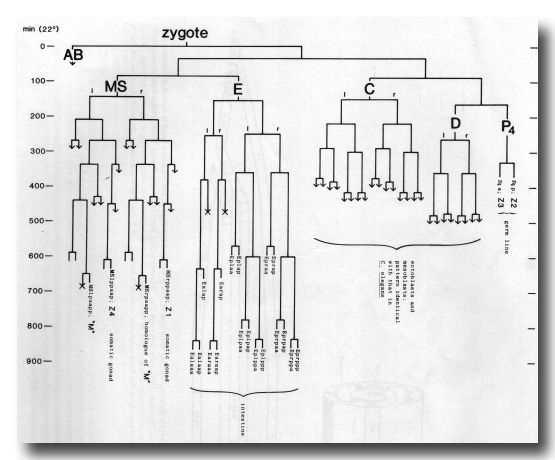 Figure 11. Partial embryonic lineage of Turbatrix aceti. Conventions as for Fig. 3. Times (at 22°C) are approximate, for reasons given in text; the time scale is adjusted to facilitate comparison with Fig. 3. Arrows represent continued division, not followed.
Figure 11. Partial embryonic lineage of Turbatrix aceti. Conventions as for Fig. 3. Times (at 22°C) are approximate, for reasons given in text; the time scale is adjusted to facilitate comparison with Fig. 3. Arrows represent continued division, not followed.
We have also followed the origin of the founder cells and certain later lineages in the embryo of Panagrellus redivivus. This nematode is of interest because Sternberg and Horvitz (1981, 1982) have shown that its postembryonic lineage is quite similar to that of C. elegans, and that the newly hatched animal contains the same set of blast cells with one addition. We find that this extra blast cell, known as T3, has the embryonic ancestry Caappa; it is therefore homologous with the neuron PVR of C. elegans. On the basis of Nomarski microscopy, PVR is absent from P. redivivus (Sternberg, personal communication). P. redivivus proved to have the same endodermal lineage as T. aceti, to which it is closely related (Ritter, 1975); in one individual, however, division of Ea(1/r)ap resulted in an intestine having 20 cells instead of the usual 18.
zur Strassen (1959) based his conclusion -that in B. rigidum P4 yields the somatic gonad- upon the observations that P4 divides in the embryo and that the newly hatched larva contains only one germ cell. Although we were not able to obtain B. rigidum, we investigated another member of the order Tylenchida, Aphelencoides blastophthorus, which similarly contains a single germ cell at hatching. We find that P4 does not divide in the embryo of A. blastophthorus, and becomes the solitary larval germ cell. Comparison with the detailed drawings provided by zur Strassen (1892) shows that the early cleavages of the two nematodes are very similar and reveals the reason for the discrepancy: he had inadvertently reversed anterior and posterior, so that the dividing cell which he took to be P4 was in fact one of the AB group.
In conclusion, although differences of detail have been seen, the general pattern of cell fates shown in figure 9 is correct for the two rhabditids C. elegans and T. aceti, and may well be conserved throughout the class Secernentea.
TISSUE DESCRIPTION
Hypodermis
The hypodermis is a sheet of cells which forms the outer surface of the nematode and secretes the cuticle. Its postembryonic development has been described by Sulston and Horvitz (1977), and other aspects have been discussed by White (1974) and by Singh and Sulston (1978), but no comprehensive account has previously been given.
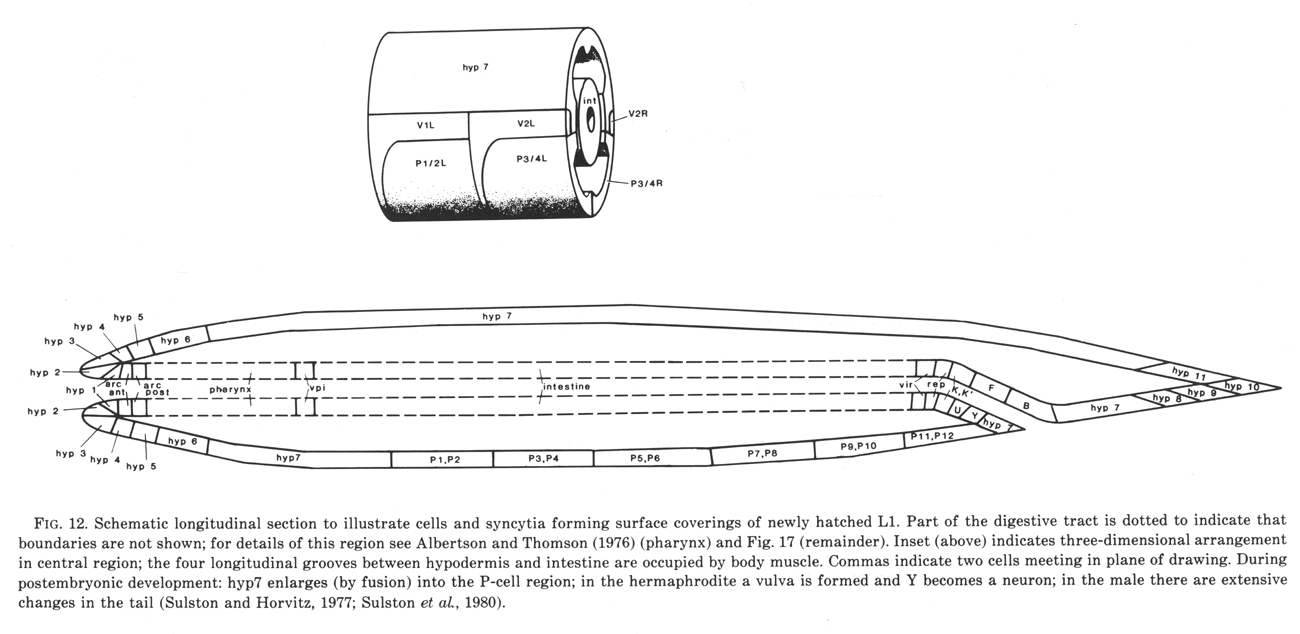
In the newly hatched L1 the anterior part of the hypodermis consists of a series of cylindrical syncytia linked together by desmosomes. These cylinders are numbered hyp1 to hyp7 from the mouth posteriorly (figure 12). The anterior and posterior arcades, which are historical names for the specialised hypodermis which lines the mouth (Chitwood and Chitwood, 1974), follow the same plan. hyp7 extends back on the dorsal side and encircles the body again at the anus; the midventral surface is occupied by the P cells (ventral cord blast cells). The tail is completed by three mononucleate cells (hyp8, 9, 11) and a binucleate cell (hyp10). All the hypodermal syncytia arise as mononucleate cells which subsequently fuse together.
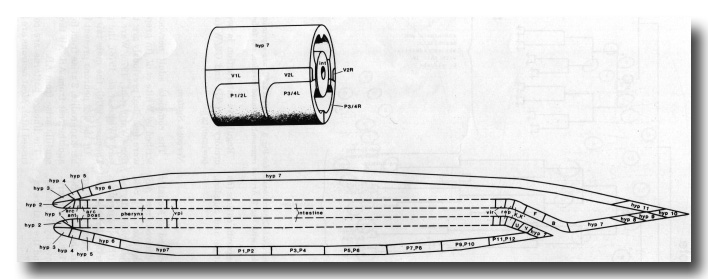 Figure 12. Schematic longitudinal section to illustrate cells and syncytia forming surface coverings of newly hatched L1. Part of the digestive tract is dotted to indicate that boundaries are not shown; for details of this region see Albertson and Thomson (1976) (pharynx) and figure 17 (remainder). Inset (above) indicates three-dimensional arrangement in central region; the four longitudinal grooves between hypodermis and intestine are occupied by body muscle. Commas indicate two cells meeting in plane of drawing. During postembryonic development: hyp7 enlarges (by fusion) into the P-cell region; in the hermaphrodite a vulva is formed and Y becomes a neuron; in the male there are extensive changes in the tail (Sulston and Horvitz, 1977; Sulston et al, 1980).
Figure 12. Schematic longitudinal section to illustrate cells and syncytia forming surface coverings of newly hatched L1. Part of the digestive tract is dotted to indicate that boundaries are not shown; for details of this region see Albertson and Thomson (1976) (pharynx) and figure 17 (remainder). Inset (above) indicates three-dimensional arrangement in central region; the four longitudinal grooves between hypodermis and intestine are occupied by body muscle. Commas indicate two cells meeting in plane of drawing. During postembryonic development: hyp7 enlarges (by fusion) into the P-cell region; in the hermaphrodite a vulva is formed and Y becomes a neuron; in the male there are extensive changes in the tail (Sulston and Horvitz, 1977; Sulston et al, 1980).
On each side of the animal there is a longitudinal row of specialised hypodermal cells, called seam cells (H0-H2, V1-V6, T); they remain separate from the rest of the hypodermis, and are responsible for making the lateral cuticular ridges known as alae (Singh and Sulston, 1978). All except HO are blast cells in the wild type, and even HO has been seen to divide in certain mutants (E. Hedgecock, personal communication).
The location of the larger hypodermal nuclei in the L1 is shown in Figs. 13 and 14. During postembryonic development many more nuclei, generated by division of the seam cells and P cells, are added to hyp7, which comes to occupy most of the body surface. For this reason hyp7 has been called the large hypodermal syncytium.
Embryonically, the hypodermis is derived from the founder cells AB and C. The dorsal nuclei undergo a strange contralateral migration in midembryogenesis (see Migrations). The finely tapering spike of the tail is formed by a process which passes posteriorly through hyp10 and contains a bundle of filaments; the process is formed by a binucleate cell (ABp(l/r)ppppppa) which subsequently dies. The dorsal ridge of hyp7 seems to act as a storage organ during late embryogenesis; the concentration of refractile granules in it becomes more and more marked while that in the intestine diminishes.
The pattern of generation of the postembryonic blast cells (nearly all of which are hypodermal at hatching) is more intriguing than informative. Most of the seam cells are made from ABarpp, but V3 and V5 are closely related to the P cells. The special origin of V5 is not unexpected, in view of its unique postembryonic programme (Sulston and Horvitz, 1977); on the other hand, there is no known difference in behaviour between V3 and the other seam cells. By ancestry, the P cells fall neatly into the equivalence groups previously revealed by postembryonic laser ablation (Sulston and White, 1980); embryonic laser ablations are consistent with the hypothesis that the equivalence groups are determined cell autonomously, although these experiments are not conclusive (see Cell Interaction Experiments).
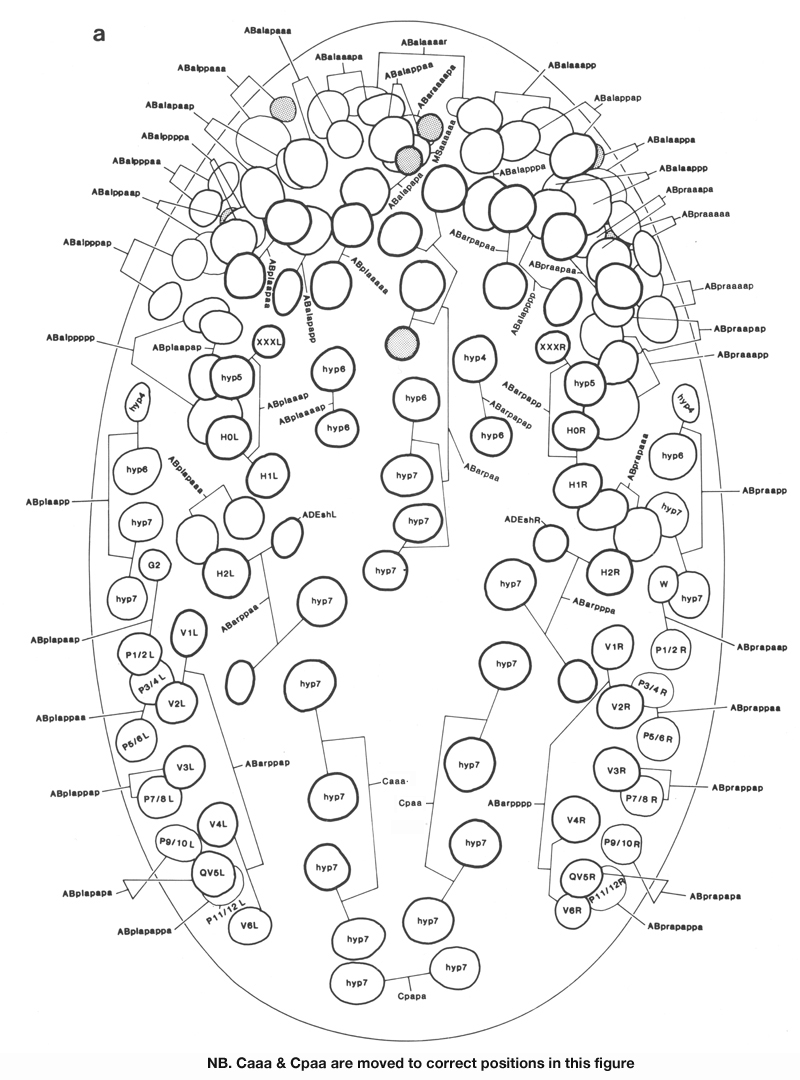
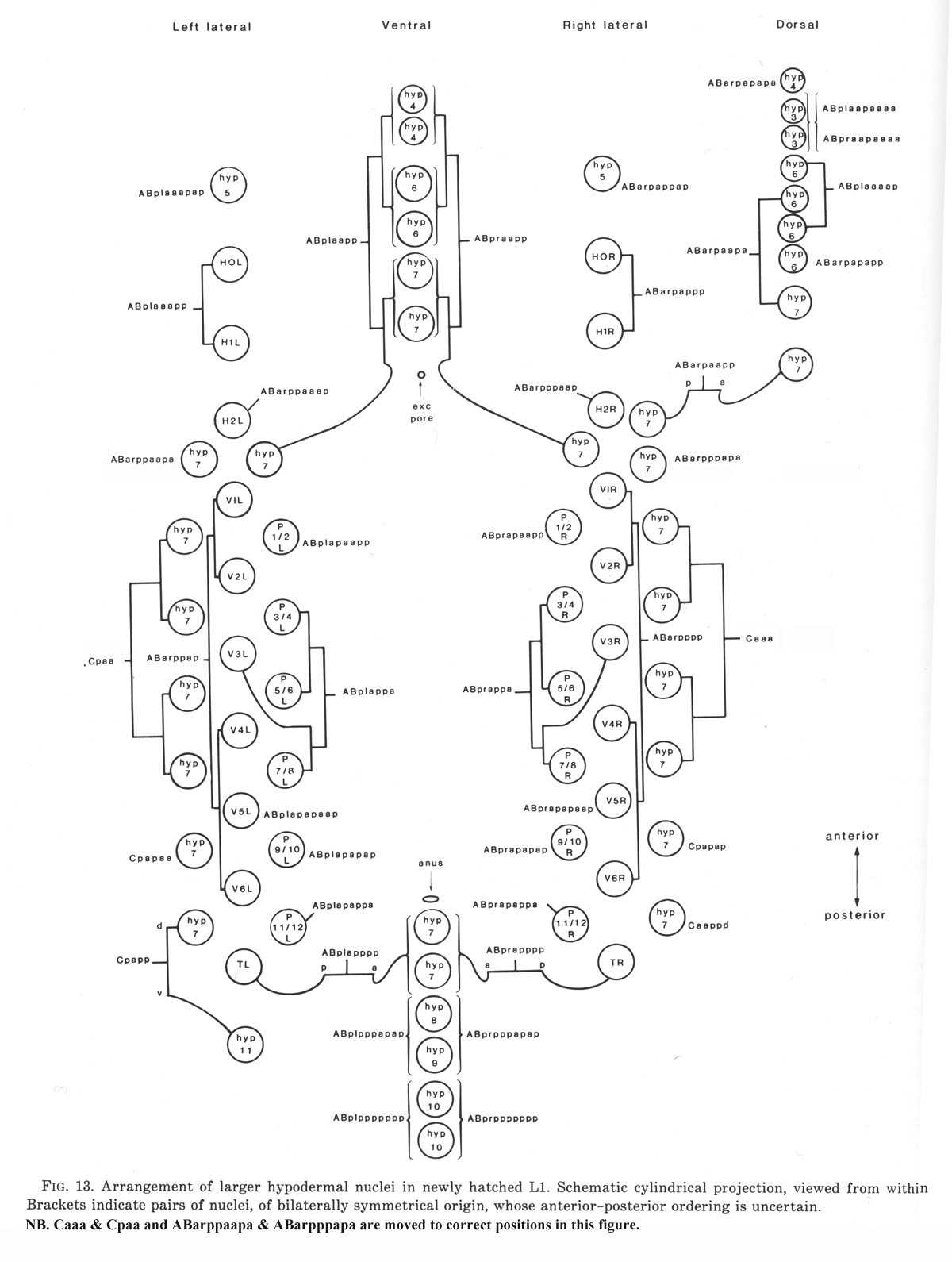
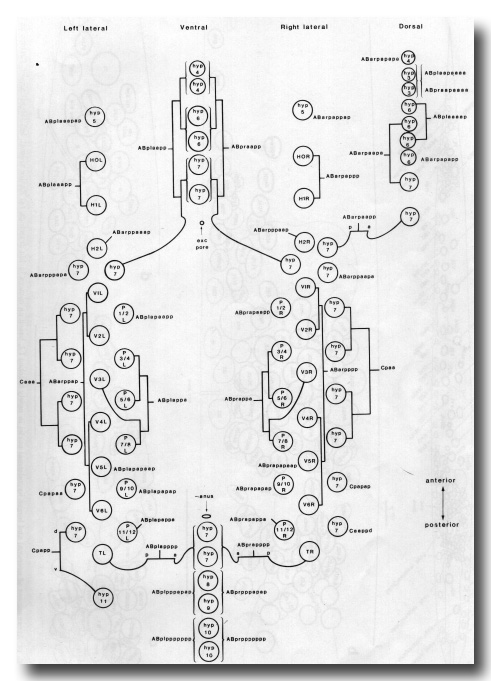 Figure 13. Arrangement of larger hypodermal nuclei in newly hatched L1. Schematic cylindrical projection, viewed from within animal. Brackets indicate pairs of nuclei, of bilaterally symmetrical origin, whose anterior-posterior ordering is uncertain. WA editors' note: the positions of the descendants of Caaa and Cpaa and ABarppaapa and ABarpppapa are inadvertantly reversed in this fig (Loer C. et al, WBG 10(3)120).
Figure 13. Arrangement of larger hypodermal nuclei in newly hatched L1. Schematic cylindrical projection, viewed from within animal. Brackets indicate pairs of nuclei, of bilaterally symmetrical origin, whose anterior-posterior ordering is uncertain. WA editors' note: the positions of the descendants of Caaa and Cpaa and ABarppaapa and ABarpppapa are inadvertantly reversed in this fig (Loer C. et al, WBG 10(3)120).
.
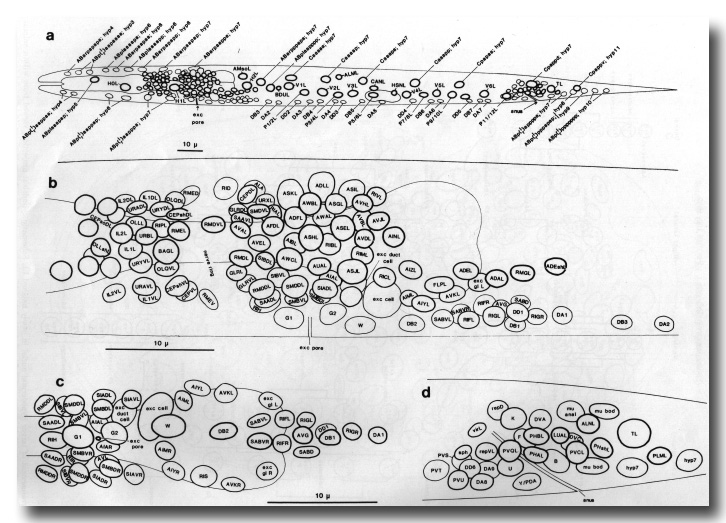 Figure 14. Arrangement of neuronal and larger hypodermal nuclei in newly hatched L1; based on camera lucida drawings. (a) Entire animal: left lateral aspect. Pattern on the right is identical, except that: additional hyp7 nucleus (ABarpaappp) lies dorsal to H2R; homologue of QL is QR, of hyp11 is PVR. (b) Ring, ventral, and retrovesicular ganglia: left lateral aspect. Note that arrangement of ring ganglion cells around posterior bulb of pharynx is very variable at this stage. Anatomy anterior to the ring is not wholly known, since cells in this region were mostly identified by their processes in the embryo. (c) Ventral and retrovesicular ganglia: ventral aspect, (d) Preanal and left lumbar ganglia, rectal cells: left lateral aspect. WA editors' note: the positions of the descendants of Caaa and Cpaa are inadvertantly reversed in this fig (Loer C. et al, WBG 10(3)120).
Figure 14. Arrangement of neuronal and larger hypodermal nuclei in newly hatched L1; based on camera lucida drawings. (a) Entire animal: left lateral aspect. Pattern on the right is identical, except that: additional hyp7 nucleus (ABarpaappp) lies dorsal to H2R; homologue of QL is QR, of hyp11 is PVR. (b) Ring, ventral, and retrovesicular ganglia: left lateral aspect. Note that arrangement of ring ganglion cells around posterior bulb of pharynx is very variable at this stage. Anatomy anterior to the ring is not wholly known, since cells in this region were mostly identified by their processes in the embryo. (c) Ventral and retrovesicular ganglia: ventral aspect, (d) Preanal and left lumbar ganglia, rectal cells: left lateral aspect. WA editors' note: the positions of the descendants of Caaa and Cpaa are inadvertantly reversed in this fig (Loer C. et al, WBG 10(3)120).
It is likely that the hypodermis is primarily responsible for the overall architecture of the animal, but the way in which it achieves this is largely unknown. One hint comes from ablation experiments in the head, which suggest that tension in the head hypodermis is necessary for elongation of the tail (see Cell Interaction Experiments). Certainly the cuticle is not involved in the shaping process, because the first sign of cuticle formation is at 600-650 min. At this time the seam cells acquire large Golgi bodies visible by Nomarski microscopy (cf Singh and Sulston, 1978), and cuticle can be seen at the mouth and in the rectum. The paired lateral alae characteristic of the L1 do not appear until just before hatching, and are apparently generated by circumferential contraction of the seam-specific cuticle (as is the case for dauer larva alae (Singh and Sulston, 1978)).
Excretory System
The excretory system has been described by Mounier (1981) and Nelson et al. (1983). It is derived from AB. Internally, it comprises four cells: the excretory cell, the duct cell, and two gland cells (which eventually become fused (Nelson et al, 1983)). A fifth cell surrounds the excretory pore, and forms the interface between the excretory duct and the hypodermal syncytia: it is known as the excretory socket or pore cell. In the embryo, the socket cell is G1; after hatching, G1 becomes a neuroblast and the socket function is taken over by G2; finally, G2 divides into a neuroblast (G2.a) and the mature excretory socket (G2.p).
Regulative interaction is seen within the pairs excretory duct/G1 and G2/W (see Cell Interaction Experiments).
Nervous System
The complete adult nervous system has been described by White et al. (in preparation). Regions which were described previously are: the anterior nervous system (Ward et al, 1975; Ware et al, 1975); the ventral cord (White et al, 1976); the pharynx (Albertson and Thomson, 1976); the male tail (Sulston et al, 1980).
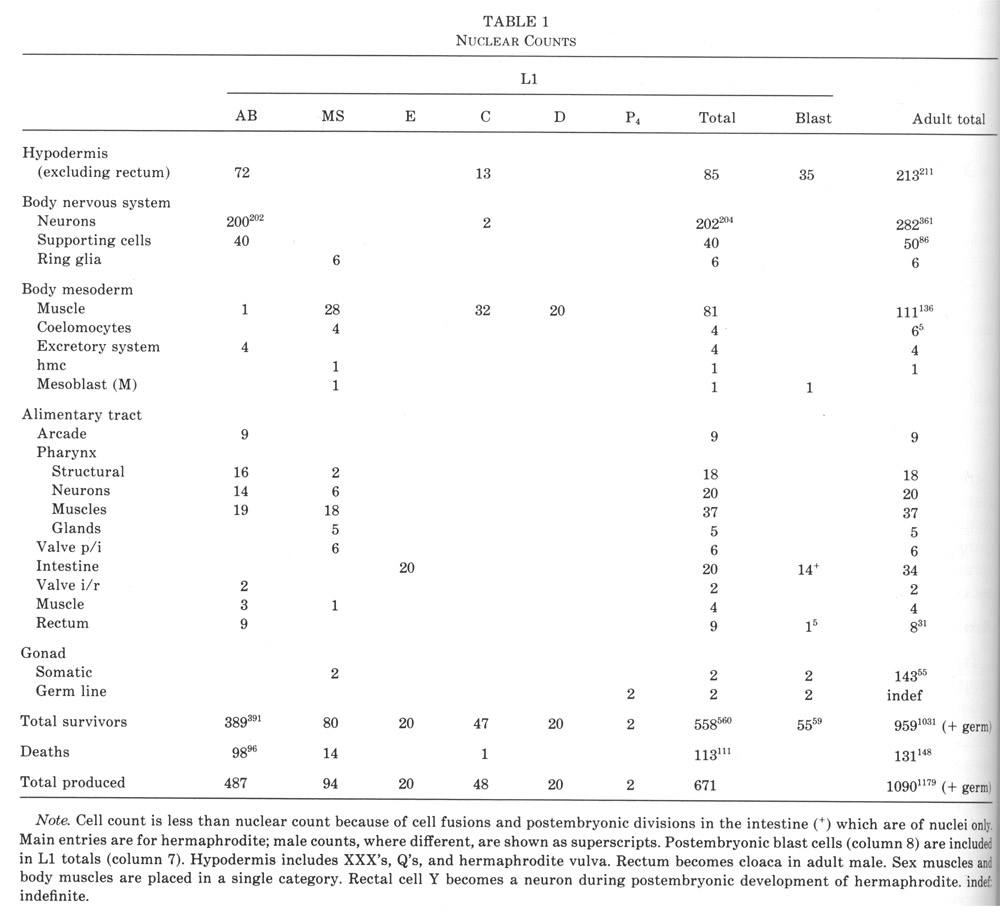
Of the 222 neurons present in the newly hatched L1 hermaphrodite, 2 arise from founder cell C, 6 from MS, and the rest from AB. All the supporting cells of the sensilla arise from AB.
The lineage patterns are complex, and will be discussed later. Most neurons are born fairly close to their ultimate positions, though a few migrate long distances (see Migrations) and many migrate short distances relative to their neighbours. There are two occasions upon which mass movements of neuroblasts and neurons are noticeable. The first (230-290 min) leads to closure of the ventral cleft at the end of gastrulation. In the second (about 400 min) the anterior neurons move towards the tip of the head, and the rudiments of the sensilla are formed; the neurons then move posteriorly again, the sensory cell bodies laying down their dendritic processes as they go. At the same time, a depression appears in the tip of the head (figure 8c); this does not involve morphogenetic cell death, and is presumably a way of providing more surface area for the sensilla. The depression is not a primordial mouth, because it subsequently everts; the buccal cavity arises further inside, between the arcade cells.
After 430 min the tip of the head elongates and the pharynx grows forward through the mass of neurons surrounding the developing nerve ring; at the same time the head becomes thinner. The pattern of neurons changes rapidly at first but stabilises after about 2 hr. In late embryogenesis it is possible to recognise all the neurons in the ring ganglion by their positions; at hatching the arrangement of the most posterior ones changes in an unpredictable way, perhaps as a result of pharyngeal movements.
At about 470 min sexual dimorphism becomes visible for the first time: in the hermaphrodite the cephalic companions (CEM) die, whilst in the male the hermaphrodite-specific neurons (HSN) die. It appears that these decisions are not made at the time that the cells are born, because all six behave at first in the same way in both sexes. In the hermaphrodite the CEMs have time to grow into the cephalic sensilla, where they form desmosomes with the sheaths and the cephalic neurons; in the male the HSNs migrate anteriorly at the same rate as they do in the hermaphrodite.
In postembryonic development a periodically repeated sublineage generates five classes of motorneurons in the ventral cord (Sulston and Horvitz, 1977; White et al, 1976). In the embryo, however, there is no such repeated sublineage to produce the three classes of juvenile motorneurons (DA, DB, and DD) which are interspersed along the ventral cord (figure 14). All that can be said is that classes DA (together with SAB, the analogue of DA in the retrovesicular ganglion) and DD are each generated semiclonally, whilst DB neurons have a variety of unique origins and are not closely related to one another.
Mesoderm (Excluding the Pharynx)
The anatomy of the larval mesoderm has been described by Sulston and Horvitz (1977).
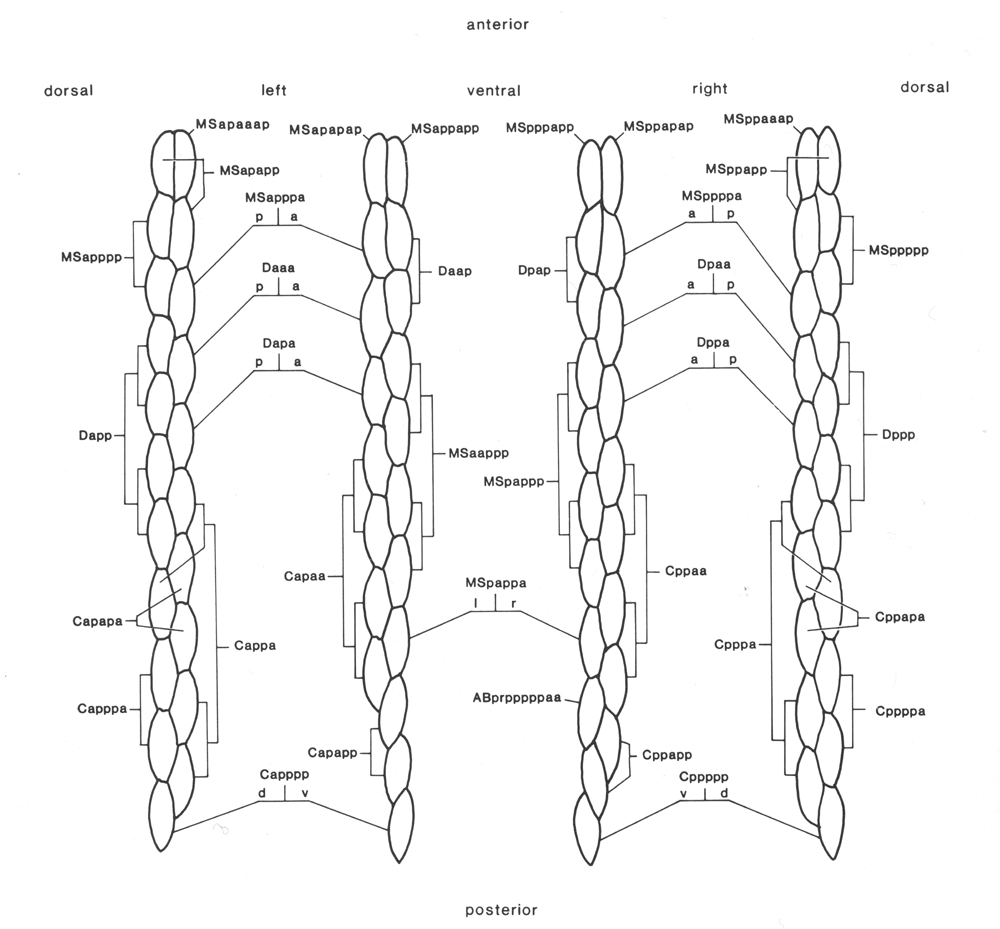
Of the 81 body muscles present in the L1, 80 are generated in a symmetrical fashion by MS, C, and D; the remaining one is generated by AB. A schematic cylindrical projection of their arrangement is shown in figure 15. The pattern of overlaps between the spindle-shaped cells is already apparent at 430 min, and allows unambiguous assignment at this stage.
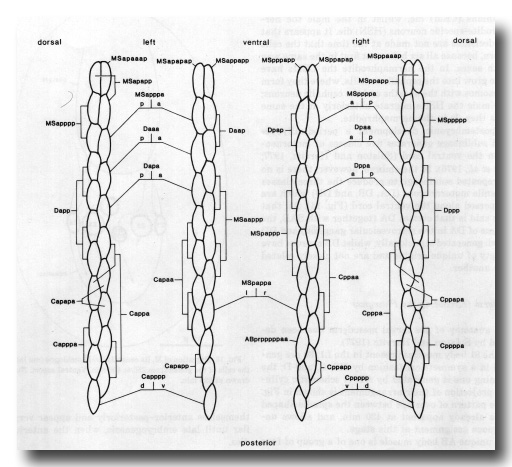 Figure 15. Arrangement of body muscles at 430 min; schematic cylindrical projection, viewed from within animal. The shapes of the muscles are not intended to be realistic, but each cell is defined uniquely by its position in the pattern. Note that, with the exception of ABprpppppaa, the assignments are bilaterally symmetrical WA editor's note: MSpappaa is shown as MSpappal in this diagram and MSpappap is shown as MSpappar (cf. Fig. 3)(R. Lee pers. comm., and Richards et al., 2013.)
Figure 15. Arrangement of body muscles at 430 min; schematic cylindrical projection, viewed from within animal. The shapes of the muscles are not intended to be realistic, but each cell is defined uniquely by its position in the pattern. Note that, with the exception of ABprpppppaa, the assignments are bilaterally symmetrical WA editor's note: MSpappaa is shown as MSpappal in this diagram and MSpappap is shown as MSpappar (cf. Fig. 3)(R. Lee pers. comm., and Richards et al., 2013.)
The unique AB body muscle is one of a group of four muscles generated preanally by AB. The two mother cells of this group (ABp(l/r)pppppa) remain on the outside of the embryo until their division at 295 min. The other three members of the group become the anal muscle, the sphincter muscle, and one of the two intestinal muscles.
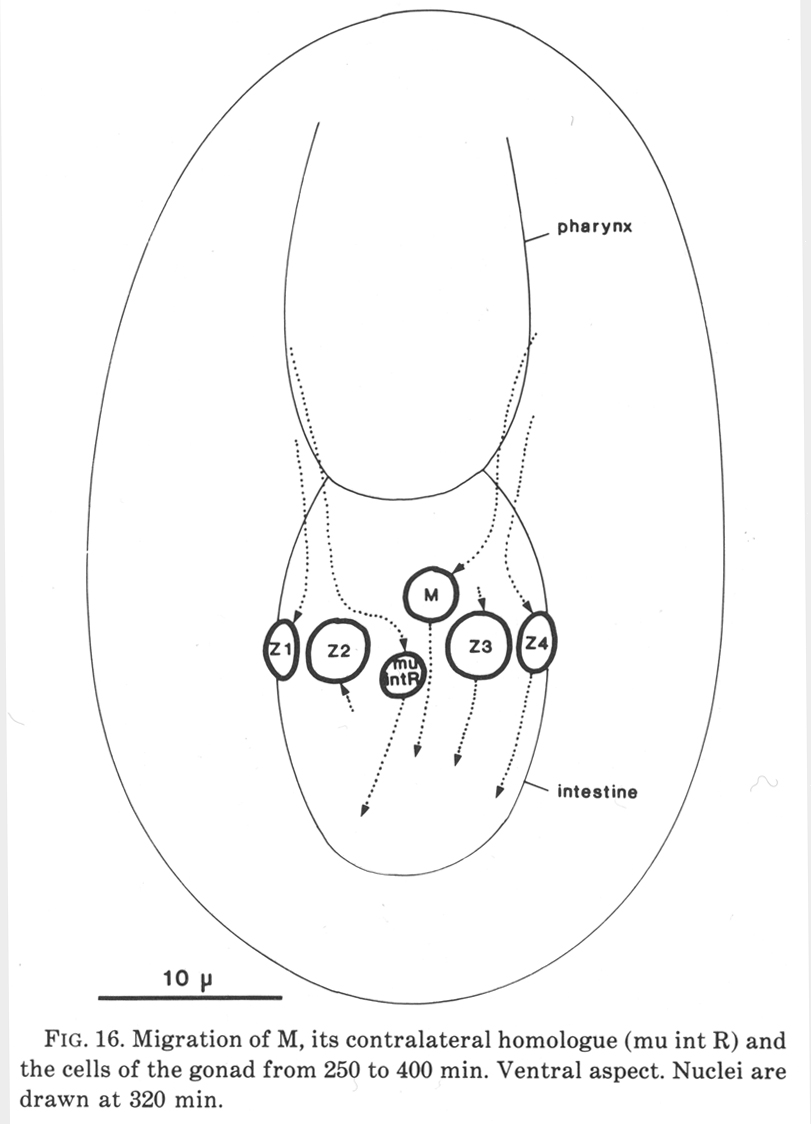
The postembryonic mesoblast (M) is born on the left, next to the pharynx. It migrates posteriorly, following a distinctive path between the two germ line cells (figure 16); it remains on the midline for some time, but then gradually shifts to the right-hand side of the intestine. The contralateral homologue of M migrates in a similar way, preceding M along the midline between the germ cells, but then differentiates into the second intestinal muscle.
The head mesodermal cell (hmc) is one of a pair of homologues (sisters to the somatic gonad cells) which migrate to the dorsal midline. There the two cells align themselves anterior-posteriorly and appear very similar until late embryogenesis, when the anterior one dies.
The four coelomocytes are generated symmetrically. Their reproducible and sexually specific arrangement arises as a result of later movements, the reasons for which are not understood.
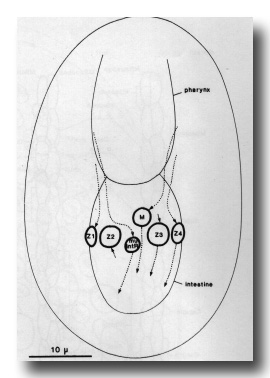 Figure 16. Migration of M, its contralateral homologue (mu int R) and the cells of the gonad from 250 to 400 min. Ventral aspect. Nuclei are drawn at 320 min.
Figure 16. Migration of M, its contralateral homologue (mu int R) and the cells of the gonad from 250 to 400 min. Ventral aspect. Nuclei are drawn at 320 min.
Alimentary Tract
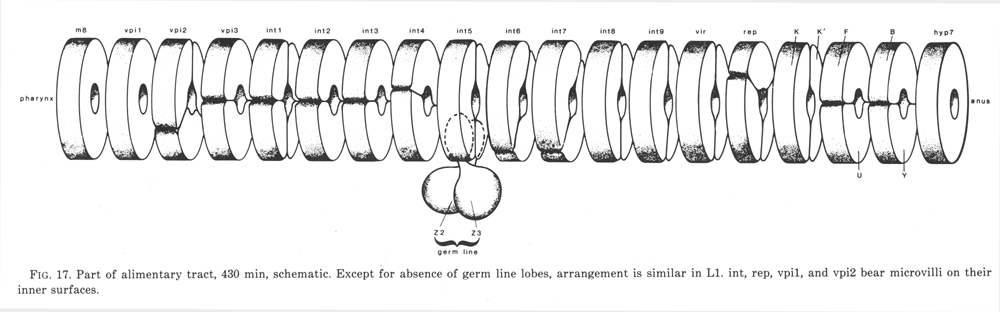
The alimentary tract is a single tube which comprises the following components: (mouth), buccal cavity, pharynx, pharyngo-intestinal valve, intestine, intestino-rectal valve, rectum, (anus). Part of it is shown schematically in figure 17.
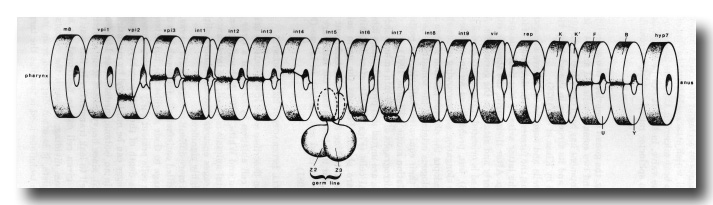 Figure 17. Part of alimentary tract, 430 min, schematic. Except for absence of germ line lobes, arrangement is similar in L1. int, rep, vpi1, and vpi2 bear microvilli on their inner surfaces.
Figure 17. Part of alimentary tract, 430 min, schematic. Except for absence of germ line lobes, arrangement is similar in L1. int, rep, vpi1, and vpi2 bear microvilli on their inner surfaces.
Buccal cavity, pharynx, and pharyngo-intestinal valve. The pharynx is a pump which ingests bacteria and crushes them; it is a complex organ, comprising muscles, structural cells, neurons, and glands. Its anatomy in the adult has been described by Albertson and Thomson (1976); their account needs amendment only in that the m2 muscles are binucleate. The buccal cavity, which is formed by the arcade cells and the anterior end of the pharynx, has been described by Wright and Thomson (1981).
All the mechanical elements of these tissues (arcade, epidermal cells, muscles, marginal cells, valve) can be recognised in the 430-min embryo by the pattern of desmosomes which they form. The neurons and glands of the pharynx were followed until they settled into the mature pattern of nuclei which persists essentially unchanged from late embryogenesis to the adult. A series of cell fusions, which take place either before or soon after hatching, yield the multinucleate cells seen in the adult. In muscle class m1, all six cells fuse together; in each of the muscle classes m2, m3, m4, and m5, the six cells fuse in pairs -DL with DR, L with VL, and R with VR; in gland class g1, AR fuses with P.
The arcade, pharynx, and pharyngo-intestinal valve are generated by two granddaughters of MS and three great-great-granddaughters of AB. These precursors, however, do not yield exclusively these tissues. At the anterior end, there are no obvious lineal boundaries between the future hypodermis, arcade, and pharynx, in spite of the specialisations which become apparent later. Conversely, there is no functional boundary between pharyngeal components derived from MS and those derived from AB. For example, apparently identical cells arise respectively from MS and AB in muscle rings m3, m4, and m5 (see Appendix), and indeed three MS muscle cells go so far as to fuse with seemingly identical AB partners (m4VR with m4R, m5VL with m5L, m5VR with m5R).
Descendants of the precursors enter the body cavity from the ventral side during late gastrulation. First to enter are the MS cells (120-200 min), next are the ABaraap cells (210 min), and last are the remaining AB cells (220-250 min). At first the dividing cells form a cylinder anterior to the intestine; gradually a distinct boundary appears at the surface of the developing pharynx; then, at about 400 min, it is compressed posteriorly and becomes almost spherical, but subsequently it gradually elongates, first anteriorly and then posteriorly. The transient compression coincides with a flux of anterior sensory neurons towards the tip of the head (see Nervous System) but the causal relationship between these events is unknown.
The three g1 gland cells migrate in a reproducible way. Their movements approximately follow the subsequent course of their secretory processes, and may be responsible for laying down the latter. The cell bodies of the anterior muscles and epidermis also move substantially in late embryogenesis (compare figure 8c with Albertson and Thomson (1976)).
The structural elements of the mature pharynx have an exact threefold rotational axis of symmetry, yet there is no trace of a threefold axis in their lineages; rather, the lineages show approximate bilateral symmetry and the third symmetry element arises by piecemeal recruitment of cells.
After 430 min (figure 8c) the pharynx continues to elongate, and within an hour the two bulbs and the isthmus are apparent. A refractile thread gradually appears along the axis of the pharynx and protrudes from the mouth. At 600-650 min the formation of L1 cuticle begins. The straight-sided cylinder of the buccal cavity appears, still plugged by the tip of the thread, and the pharyngeal lumen becomes outlined. One hour before hatching, the g1 glands become active, just as they do before ecdysis (Singh and Sulston, 1978). Half an hour later the pharynx begins to pump spasmodically, the mouth plug falls away, and the refractile thread is broken up and discharged into the intestine.
Intestine.The intestine comprises a chain of paired cells (figure 17). At hatching they are mononucleate, but subsequently most of them become binucleate by nuclear division (Sulston and Horvitz, 1977). Occasionally an extra cell is found in a newly hatched larva, presumably as a result of an extra division in the E lineage (cf. Other Nematode Species: P. redivivus). The anterior ring of four cells (int1) is specialised in having shorter microvilli than the rest of the intestine.
The intestine is derived exclusively from founder cell E, which gives rise to no other tissue. The daughters of E are the first cells to enter the body cavity during gastrulation (90 min). By 300 min they have formed two rows of eight cells, one on the left and one on the right. The anterior pair divide dorso-ventrally to yield int1, which attaches to the pharyngo-intestinal valve. Before attaching to int1, the rest of the anterior intestine undergoes a 90° left-handed twist, so providing half the total twist noted by Sulston and Horvitz (1977); the remainder seems to be due to packing of the posterior nuclei, because no twist is seen in the attachment of the intestine to the intestino-rectal valve (Fig. 17).
Intestino-rectal valve and rectum. The cells which form these structures are shown schematically in figure 17. Some of them have been given new names: rectal epithelium was formerly rectal gland, U was E, and Y was C. K is a blast cell; its contralateral homologue K' is a blast cell in the C. elegans mutant lin-4 (Chalfie et al, 1981) and also in Panagrellus redivivus (Sternberg and Horvitz, 1982). F, U, B, and Y are blast cells in the male. All these cells underlie cuticle in the L1. During late larval development of the hermaphrodite Y withdraws from the hypodermis, without division, and becomes a neuron.
There is some similarity between the intestino-rectal valve and the pharyngo-intestinal valve: in both, the intestine attaches to a ring of two cells which do not bear microvilli, which attaches in turn to a ring of three cells which do bear microvilli. Only the intestino-rectal valve is a true valve, in that it can be closed actively by means of a sphincter muscle which surrounds it.
Gonad
At hatching, the gonad comprises two germ line cells (Z2 and Z3) and two somatic cells (Z1 and Z4). Its subsequent development has been described by Kimble and Hirsh (1979), and its structure in the adult by Hirsh et al. (1976) and by Klass et al (1976).
The germ cells and somatic cells have separate embryonic origins; the former are the daughters of founder cell P4, and the latter arise by identical lineages from MSa and MSp. After their birth, the somatic cells migrate posteriorly and attach to the germ cells. At this stage the gonad is oriented transversely across the animal (figure 16), but, probably as an inevitable consequence of the elongation and narrowing of the embryo, it gradually adopts the oblique position shown in figure 8c. The homologous origin of the somatic cells is concordant with their equivalent behaviour in Postembryonic laser ablation studies (Kimble, 1981).
Electron micrographs of a 470-min embryo show that the germ cells are united and protrude large lobes into two intestinal cells (Figs. 2j, 17); after hatching, the protrusions are absent. Perhaps the germ cells are nursed by the intestine until their attachment to the somatic cells, for it is known that the latter are essential for their survival and division in larvae (Kimble and White, 1981).
CELL INTERACTION EXPERIMENTS
This section describes some investigations using the technique of cell ablation by means of a laser micro-beam (Sulston and White, 1980). Although the number of experimental animals is small, the invariance of the wild-type lineages ensures that any abnormalities observed are highly significant.
Ablation is more difficult in eggs than in larvae. Small cells can be killed satisfactorily (although it is difficult to avoid damage to their neighbours), but attempts to kill the large cells present early in embryogenesis frequently cause death of the entire embryo. Damage to these young eggs can be minimised by mounting them on 1% agar in an isotonic medium (see Light Microscopy: T. aceti), since heavy pulses seem sometimes to cause the shell to leak transiently. In this way the nucleus of any given cell can be destroyed, or at least prevented from dividing; although the cytoplasm inevitably persists, it is often displaced from its usual position. The technique used is to pulse the target cell repeatedly at an energy level sufficient to produce refractile debris in its nucleus; whenever the cell appears dangerously weakened (low refractility of the cytoplasm, excessive Brownian motion) it is rested for a minute or two before pulsing is continued. After a successful operation the cell, heavily loaded with refractile debris, appears largely to have lost contact with its neighbours and remains visible as an undivided blob.
Early Ablations
After ablation of P1, AB continued to cleave and in due course generated a large number of cell deaths. There was no recognisable organogenesis or morphogenesis, and no particular cells could be identified by light microscopy, though nuclei characteristic of hypodermal and neural tissue were visible.
In a series of animals, the two daughters and four granddaughters of AB were ablated in turn. The remaining cells divided in a superficially normal way, and intestinal, dorsal hypodermal, and germ line cells became recognisable. The arrangement of tissues, however, was defective, and all these embryos arrested at the comma stage (see figure 4).
Ablation of P4 led to the production of larvae with no germ line. The somatic gonads of these animals grow more or less normally but are devoid of gametes: P4 is not replaced by any other cell.
Although these results are superficially consistent with cell autonomous development, they are inconclusive because the presence of the dying cells may inhibit potential replacements (see Discussion).
Body Muscle from C and D
The following cells were ablated in a series of animals: Da, Dp, Capa, Capp, Cppa, Cppp. In each case, the resultant larva lacked approximately the quota of muscles which the dead precursor would normally have made (accurate muscle counts, especially in damaged animals, are difficult). The missing cells left a gap, and the remaining muscles did not spread out much to fill it. The experiments are subject to the caveat given for the early ablations.
MS Lineage
A number of experiments were carried out at the MS4 to MS16 stages. The great advantage of this group of cells is that they are gastrulating at the time of ablation; in favourable cases, the target cell can be damaged sufficiently to cause it to detach from its neighbours and to remain at the surface of the embryo for some time, although eventually it, too, becomes enclosed within the body cavity. This circumstance should permit the remaining cells to interact regulatively, if they are capable of doing so.
Various derivatives of MS(p)p were ablated at MS4 or MS8, and the following cells were scored in the L1 and later larval stages: Postembryonic mesoblast (M), right intestinal muscle (homologue of M), coelomocytes, somatic gonad (Z1, Z4: regarded as equivalent), head mesodermal cell (difficult), body muscle complement (approximate). In all cases the survivors appeared to generate those cells, and only those cells, which they would have produced in an intact embryo.
This result is particularly striking in the case of M and the right intestinal muscle, which are homologous and migrate along the midline in contact with one another (figure 16). M is a large cell with a characteristic Postembryonic lineage; it is therefore easy to score, and the result is very clear-cut (five animals). This experiment and the following one are important counterexamples to the regulative interaction shown by two pairs of AB cells (see below).
The head mesodermal cell (hmc) is more difficult to score, so the finding in this case was confirmed by watching for the death of its homologue. When MSapp or MSappa was ablated (three animals) the death was seen and hmc was absent from the larva; when MSppp or MSpppa was ablated (three animals) the death was not seen and hmc was present in the larva. Therefore, in spite of the similar embryonic appearance and position of hmc and its homologue, the latter is programmed to die even in the absence of the former.
Extension of these experiments to MS(a/p) leads to pharyngeal damage, as predicted from the lineage. Not surprisingly, these animals feed poorly or not at all. Detailed analysis, which would require electron microscopical reconstruction, has not been undertaken.
AB Lineage
In the first two sections below, experiments which produced viable animals are described. In (a) the resulting animals were scored solely by light microscopy; in (b) the ultrastructure of the anterior sensilla was determined by electron microscopy. A third section (c) lists some experiments in which the animals died.
(a) The following cells were ablated at the AB32 stage: ABplapa (three animals), ABplaaa (one animal), ABarppa (one animal). The resulting larvae were scored, by Nomarski microscopy, for the postembryonic blast cells and certain neurons; although the blast cells were sometimes displaced in experimental animals they could still be accurately identified by their division patterns. With one exception (see below), the only blast cells missing were those normally generated by the ablated precursor. Furthermore, the surviving blast cells behaved normally during Postembryonic development, even to the extent of respecting the equivalence group boundaries (Sulston and White, 1980) in the usual way.
Because of the presence of the dying AB cells on the surface of the embryos these results do not unambiguously demonstrate cell autonomy, though they are consistent with it. The best evidence is that from ABplapa, in that the progeny of this precursor would normally move to two separated regions in the embryo, and it is unlikely that a dying blob of cytoplasm could make appropriate contacts in both places.
The exception was that, after the ablation of ABplapa, W was missing and G2 was present. In these animals ABprapaapa, which normally becomes W, presumably became G2 in place of the missing ABplapaapa. This example of regulative interaction was confirmed by the direct ablation of ABplapaapa (one animal) and ABprapaapa (two animals) at 260 min, before the cells moved ventrally: all three animals subsequently formed G2 and lacked W.
A second example of regulation was revealed by the ablation of ABplpaaaap (two animals) and ABprpaaaap (one animal): all three animals subsequently formed the excretory duct (normally ABplpaaaapa) and lacked G1 (normally ABprpaaaapa).
It appears that G2/W and duct/G1 are a forward extension of the P-cell regulative system. The latter is a series of six pairs of lateral cells (Fig. 13), which move to the ventral midline during postembryonic development. The members of each pair are equivalent, but in certain cases they compete for a particular "primary" fate (Sulston and White, 1980). The presumptive G2/W and duct/G1 move to the ventral midline and compete for a primary fate in the same way but at an earlier time.
(b) The following cells were ablated at the AB32 stage: ABalaaa, ABalaap, ABalapa, ABalapp, ABalppa, ABalppp, ABpraaa, ABprpaa. The pattern of cell deaths was scored at 270 min, certain neurons (RMED, ALA, RID, CAN) and the postembryonic blast cells were scored in the young L1, and finally the anterior sensilla of the L4 larva or adult were reconstructed by serial section electron microscopy. The results concerning the anterior sensilla are collected in Fig. 18.
In each case, those components which would normally have been generated by the ablated precursor were not seen in the resulting animal, indicating that no replacement of one cell by another had occurred (figure 18). This result strongly suggests that the selection of asymmetrical precursors for symmetrical roles in the head (see Discussion: Sublineages) is a cell autonomous process, because, according to the lineage, the time of selection is mostly later than the time of ablation: in this interval the dying cell acquires a position and a shape very different from that normally occupied by its progeny.
 Figure 18. Laser ablations of precursors of anterior sensilla. Intact. Anterior sensilla are represented by ovals, enclosing symbols representing neurons and supporting cells (see Ward et al, 1975, for realistic representation). In inner labials (IL), triangle represents two neurons. In amphids (AM), "wing" cells (AWA, AWB, AWC) are not scored; remaining amphid neurons are split into three groups: AFD ("finger" cell, scorable), a group of five cells sending seven ciliated processes, and a group of three cells sending three ciliated processes into the amphid channel (the two neurons with double processes -ADF and ADL- are included in the seven process group, but otherwise the designations are purely numerical. Ablations. Ablated precursor is shown at top of each figure. Empty symbols indicate that the designated cell was not found; stippled symbols indicate uncertainty; X's indicate the cells which the ablated precursor would normally have produced. Note that, without exception, X's cover empty symbols. In some animals neurons became associated with inappropriate sockets; since sockets tended to remain in their usual positions, these changes are indicated by arrows leading from the neurons (and their sheaths: see below) to the sockets. Criteria used to identify particular cell types included: striated rootlets in IL1 and OLQ neurons; square linked array of four filled microtubules in OLQ neurons; dark deposit and supernumerary microtubules at tip of CEP neurons; prominent lips generated by Il sockets. In the region reconstructed there are no criteria for distinguishing between types of sheath; where a neuron is transferred to a different socket, so that the sheath designation is ambiguous, the Vernon is arbitrarily assumed to move with its own sheath.
Figure 18. Laser ablations of precursors of anterior sensilla. Intact. Anterior sensilla are represented by ovals, enclosing symbols representing neurons and supporting cells (see Ward et al, 1975, for realistic representation). In inner labials (IL), triangle represents two neurons. In amphids (AM), "wing" cells (AWA, AWB, AWC) are not scored; remaining amphid neurons are split into three groups: AFD ("finger" cell, scorable), a group of five cells sending seven ciliated processes, and a group of three cells sending three ciliated processes into the amphid channel (the two neurons with double processes -ADF and ADL- are included in the seven process group, but otherwise the designations are purely numerical. Ablations. Ablated precursor is shown at top of each figure. Empty symbols indicate that the designated cell was not found; stippled symbols indicate uncertainty; X's indicate the cells which the ablated precursor would normally have produced. Note that, without exception, X's cover empty symbols. In some animals neurons became associated with inappropriate sockets; since sockets tended to remain in their usual positions, these changes are indicated by arrows leading from the neurons (and their sheaths: see below) to the sockets. Criteria used to identify particular cell types included: striated rootlets in IL1 and OLQ neurons; square linked array of four filled microtubules in OLQ neurons; dark deposit and supernumerary microtubules at tip of CEP neurons; prominent lips generated by Il sockets. In the region reconstructed there are no criteria for distinguishing between types of sheath; where a neuron is transferred to a different socket, so that the sheath designation is ambiguous, the Vernon is arbitrarily assumed to move with its own sheath.
In most cases there were a few unpredicted deficiencies in the sensilla. These can be accounted for in various ways: the difficulty of recognising supporting cells in the absence of their neurons, displacement of unsupported neurons beyond the zone of reconstruction (in several cases such neurons were found lying beneath the hypodermis and well posterior of their usual positions), and perhaps failure of expression as a result of displacement. Since these missing cells are not numerous, they do not seriously weaken our conclusion.
Although replacement regulation at the cellular level was not seen, these experiments provided some interesting examples of anomalous assembly of sensilla (see arrows in figure 18). One clear conclusion is that socket cells are not restricted to association with neurons of a particular type.
(c) A number of ablations in the AB lineage repeatedly yielded animals that failed to hatch. Many of the precursors for which this was true (ABplaaa (two animals), ABarpap (five animals), ABpraap (two animals), ABplapp (seven animals), ABprapp (one animal) are normally responsible for generating large patches of hypodermis. After ablation of any one of them, overall morphogenesis proceeded as usual until the start of elongation at 400-450 min, and then the cells inside the embryo oozed out through the missing area. Elongation ceased when this happened, suggesting that it is driven by circumferential contraction in the anterior part of the embryo.
Ablation of ABplpap (three animals) and ABprpap (three animals) also caused problems. Of these animals, only one of the former hatched, apparently because the dying precursor interferes with closure of the ventral cleft. However, before the embryos burst, it was possible to see that the excretory cell was made only when ABplpap was present. The one animal that hatched lacked both the excretory cell and the rectum, as predicted, and failed to grow.
Other Late Ablations
Various other cells were ablated at 250-300 min. The object of these experiments was primarily to deduce the function of the cells or of their progeny (e.g., interneurons of the ventral cord, rectal cells, postembryonic blast cells). No further replacement regulation was revealed, but many cells remain untested. Three examples of mosaic assembly of the nematode are of interest: after ablation of ABplpapppa (parent of K and K', figure 17) the rectum was blocked; after ablation of ABprpapppp (parent of the intestino-rectal valve cells, figure 17) the posterior end of the intestine was not connected to the rectum; after ablation of ABalpaappp and ABalpapaaa (parents of hyp2) the buccal cavity was formed in the normal way at the anterior end of the pharynx, but was not connected to the mouth.
Summary
Two cases of regulative interaction were found. Both occur in late embryogenesis and involve the confrontation of similar cells at the ventral midline, a situation which is already familiar from Postembryonic studies. In other respects the results are consistent with cell autonomous development, the evidence for autonomy being strong for the MS and late AB lineages. Interactions before the 50-cell stage are difficult to analyse by laser ablation.
Discussion
Invariance, Cell Autonomy and Cell Interaction
Blastomere isolation
Cell ablation
Early divisions of the founder cells
Summary
Embryonic Germ Layers and Cell Fate
Lineal Boundaries and Functional Boundaries
Segments
Sublineages
Programmed Cell Death and Sexual Dimorphism
Rotational Symmetry
Twofold
Threefold
Four- and sixfold
Symmetry and Asymmetry
Conclusion
In this paper we have described the cell lineage that gives rise to the newly hatched larva of Caenorhabditis elegans. Since postembryonic development has already been followed (see Background Information for references), our knowledge of the cell lineage of this organism is now complete. The embryonic cell lineage is drawn out in Fig. 3, a complete list of cells is given in the Appendix, and a summary of cell types appears in Table 1.
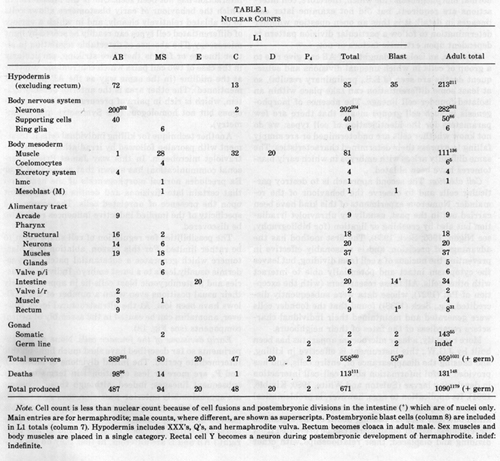 Table 1. Note. Cell count is less than nuclear count because of cell fusions and Postembryonic divisions in the intestine (+) which are of nuclei only. Main entries are for hermaphrodite; male counts, where different, are shown as superscripts. Postembryonic blast cells (column 8) are included in L1 totals (column 7). Hypodermis includes XXX's, Q's, and hermaphrodite vulva. Rectum becomes cloaca in adult male. Sex muscles and body muscles are placed in a single category. Rectal cell Y becomes a neuron during Postembryonic development of hermaphrodite, indef: indefinite.
Table 1. Note. Cell count is less than nuclear count because of cell fusions and Postembryonic divisions in the intestine (+) which are of nuclei only. Main entries are for hermaphrodite; male counts, where different, are shown as superscripts. Postembryonic blast cells (column 8) are included in L1 totals (column 7). Hypodermis includes XXX's, Q's, and hermaphrodite vulva. Rectum becomes cloaca in adult male. Sex muscles and body muscles are placed in a single category. Rectal cell Y becomes a neuron during Postembryonic development of hermaphrodite, indef: indefinite.
The main value of this information is as a basis for more detailed studies of development, though certain features of it seem of more general interest and are discussed below. There is, however, little point in thinking about the details of the cell lineage unless it has some function beyond mere cell proliferation; we shall therefore ask at the outset the question: To what extent are the fates of cells constrained by their ancestry?
Invariance, Cell Autonomy, and Cell Interaction
The embryonic cell lineage is essentially invariant. The patterns of division, programmed death, and terminal differentiation are constant from one individual to another, and no great differences are seen in timing. At any given moment in development, each blastomere not only has a predictable future but also has a reproducible position and a defined group of neighbours.
Although invariance is consistent with cell autonomous development, it is not in itself diagnostic, because it might be the result of either the absence or the reproducibility of cell-cell interactions; in order to distinguish between these possibilities it is necessary to perturb the system experimentally. Two approaches have been fruitful.
Blastomere isolation. The first approach is the isolation of early blastomeres in a supportive medium and the observation of their subsequent development. Using this method, Laufer et al. (1980) demonstrated that the unique characters of the founder cells (see General Description) are internally determined and do not depend upon interaction with neighbouring cells. They found that the progeny of isolated blastomeres do not undergo normal morphogenesis (for which, therefore, cell interactions are required), but did not examine later cell lineages in detail; it is thus an open question whether determination to follow a particular division pattern is dependent upon extrinsic factors or not.
However, an isolated daughter of AB can give rise to a group of cells in which unequal divisions and subsequent deaths are seen (J.E.S., preliminary results), so at least some differentiation can take place within an isolated founder cell lineage. The absence of morphogenesis in such cell groups means that there are few parameters for the identification of cell types: we do not know whether cells are undetermined or are simply failing to express their determined characteristics. The same difficulty arises with embryos in which early blastomeres have been ablated.
Cell ablation. The second approach is to destroy particular cells and to observe the behaviour of the remainder. Numerous experiments of this kind have been carried out in the past, usually by ultraviolet irradiation but also by crushing or ligation (for bibliography, see Nigon, 1965; Seek 1938). The first method has the advantage of precision, and is reasonably effective in preventing the nucleus of a cell from dividing, but leaves the cytoplasm intact and potentially able to interact with other cells. All these researchers (with the exception of Pai (1927), whose data were subsequently discredited, e.g., Seek (1938)) found that the founder cells were generated and maintained their individual characters regardless of the fates of their neighbours.
More recently, a laser microbeam apparatus has been built by J.G.W.; this instrument is effective in killing, and causing the disappearance of, entire cells, and has provided useful information about cell-cell interaction in developing larvae (Sulston and White, 1980; Kimble, 1981). Its application to eggs, however, is problematical because, except at relatively late stages, cells cannot be killed outright without causing death of the organism. Nevertheless, earlier blastomeres can be prevented from dividing and displaced; their relationships with other cells are thereby presumably altered, and the presence of an abnormally positioned large blob of cytoplasm probably distorts the relationships between the surviving cells as well. In this way a reasonable test for cell-cell interaction can be applied from about the 50-cell stage onwards. In a series of experiments covering this period, no examples of cell-cell interaction affecting embryonic lineage, and only two examples of interaction affecting fate, were found. As explained above (Cell Interaction Experiments: AB lineage), the latter two cases can properly be regarded as part of a postembryonic regulative episode in which pairs of homologous cells confront one another at the midline.
It is worth emphasising two areas in which regulative interaction has not been seen. One is the mesoderm, in which the behaviour of early blastomeres allows cells to be ablated relatively cleanly, and in which a variety of differentiated cell types can readily be scored by light microscopy. The absence of detectable regulation in either lineage or fate is therefore striking, particularly in the case of two cell pairs which confront one another at the midline (in the same way as the AB pairs just mentioned). The other area is the anterior sensory system, which is rich in pairs of precursors that are analogous but not homologous (see Symmetry and Asymmetry).
Another technique for killing individual cells is treatment with psoralen followed by irradiation with an ultraviolet microbeam. In this way James Priess (personal communication) has shown that removal of E or Ea precludes normal morphogenesis of the head, and that certain late divisions and deaths are dependent upon the presence of unrelated cells. The source and specificity of the implied inductive influences remain to be discovered.
The possibilities for regulation of cell shape seem to be rather limited. For this reason, ablation of a blastomere which generates a substantial patch of hypodermis usually leads to a burst embryo. Individual muscles and Postembryonic blast cells lie in approximately their usual positions even when a number of their fellows have been lost. At the ultrastructural level, however, anomalies can be caused in the assembly of neural components (see Fig. 18).
Early divisions of the founder cells. None of the experiments so far described have shed much light on this intermediate period. The initial divisions of MS, C, D, and P4 are more or less equational in terms of their subsequent lineages; indeed, although these divisions are approximately anterior-posterior, they give rise ultimately to left-right symmetrical groups of cells. (This pattern is more apparent in Turbatrix and Ascaris, because the initial divisions of MS, C, and D are left-right.) AB and E are different, in that they do not generate elements of left-right symmetry until their second rounds of division. Nevertheless, their initial divisions may be equational. In particular, the first anaphase of AB is transverse across the midline, and only at telophase do its daughters skew into an anterior-posterior configuration; this progression is seen more clearly in Ascaris, in which the transverse arrangement persists for much longer.
Classical attempts to investigate the cell autonomy of the early AB lineage in Ascaris, by the study of squashed eggs and giant eggs, led only to a mass of circumstantial data and heated controversy (see Seck, 1938). Blastomere isolation experiments would seem to offer the best hope of settling the matter, though this approach runs into further difficulties, as explained above. For the present, the bases for the difference between ABa/ABp, as well as those for the more subtle distinctions between MSa/MSp, Ea/Ep, and Ca/Cp, remain unknown.
Summary. In our limited survey, no replacement of any embryonic precursor by another was observed, but the postembryonic behaviour of two pairs of cells was found to be decided by regulative interaction in midembryogenesis. There is evidence for inductive effects upon morphogenesis, cell divisions, and cell deaths during the second half of embryogenesis. The period between the generation of the founder cells and the 50-cell stage has not yet been satisfactorily investigated.
Embryonic Germ Layers and Cell Fate
In their literal sense, the terms ectoderm, mesoderm, and endoderm refer to the three distinct layers of cells formed at gastrulation in triploblastic embryos -the so-called germ layers. By extension, the same terms are applied, in mature animals, to the groups of tissues typically derived from the germ layers. To avoid confusion, we shall qualify the terms with "embryonic" when using them in the former sense.
It has long been known that the correlation between germ layer and tissue type is not perfect. In vertebrate embryos, for example, part of the embryonic ectoderm, known as the neural crest, contributes cells to structures (such as muscle and cartilage) which are more typically derived from embryonic mesoderm (reviewed by le Douarin, 1980). Is this also true for the nematode?
The ancestry of various cell types in C. elegans is summarised in Fig. 9. From the behaviour of their progeny at gastrulation the founder cells can be classified as follows: -embryonic endoderm: E; embryonic mesoderm: MS, D, part of C; embryonic ectoderm: AB, remainder of C (though the progeny of AB which contribute to the pharynx might also be classified as embryonic mesoderm). Thus neurons can be derived from embryonic mesoderm and muscles from embryonic ectoderm.
The mingling in ancestry is most extensive in the pharynx. For example, there are three cases of neurons which are sisters to muscles (strictly, myo-epithelial cells), showing that divergence between these two fates can occur as late as the terminal division. Conversely, a consideration of symmetry and patterns of cell fusion in muscle rings m3, m4, and m5 (see Alimentary Tract and Appendix) indicates that a particular cell type may be generated by more than one developmental pathway.
The pharynx may perhaps be regarded as atypical, in that it is a remarkably self-contained organ with its own nervous system and highly specialised myo-epithelial cells. The very late derivation of four muscle cells from the AB lineage is a more straightforward example. The parents of these cells remain on the outside of the embryo until 290 min. Two of the muscles are associated with the rectum and are unique, but the other two are identical with cells derived from MS: one is an intestinal muscle and the other is a body muscle.
The conclusion is that, although there is a broad mapping of particular cell types onto particular blast cells, absolute distinctions are not laid down at an early stage. One reasonable explanation for this observation is that the broad categories correspond to a primitive ancestral condition, and that in the course of evolution their boundaries have occasionally been transgressed; such an event may well be improbable, in that it would involve reprogramming a partially committed cell, but may nevertheless occasionally be selected.
Lineal Boundaries and Functional Boundaries
Two identifiable tissues are generated as single exclusive clones; i.e. in each case one precursor generates all the cells of the tissue and no others. These tissues are the intestine (founder cell E) and the germ line (founder cell P4).
Such coincidence of lineal boundaries with functional boundaries is the exception rather than the rule. Examples of partial clonal derivation can be recognised: some precursors generate only, but not all, cells of a given type (e.g., D, Cap, and MSpppp all yield exclusive clones of body muscle); other precursors generate all, but not only, cells of a given type (e.g., MS yields all the coelomocytes and the entire somatic gonad). However, throughout most of the lineage cell types are intermingled even at the terminal divisions.
As a result of this heterogeneity of ancestry with respect to cell fate, many lineal boundaries cross the various somatic structures in an apparently meaningless way. For example, the neurons and supporting cells of a given embryonic sensillum never arise as an exclusive clone, and the boundary between AB and MS meanders through binucleate cells.
It is interesting that exactly the same conclusion is reached with regard to polyclones in the epidermis of insects (Garcia-Bellido et al, 1973; Lawrence, 1981). Here, the reproducible lineal boundaries surround the progeny not of single cells but of groups of cells called compartments; the cell lineages within a given compartment are indeterminate. Like the clonal boundaries in the nematode, compartment boundaries do not necessarily circumscribe recognisable functional units and yet are reproducibly positioned from one individual to another.
Segments
Nematodes are not usually considered to be segmented animals. However, the lateral hypodermis of the newly hatched L1 larva is divided into periodically repeated units (Fig. 13). The repeat unit is most obvious in the middle of the body, where it comprises a ventral cord blast cell (P), a lateral hypodermal blast cell (V), and a syncytial hypodermal nucleus (hyp7); the head and the tail can plausibly be regarded as containing additional degenerate units. How do these units arise?
Inspection of the embryonic lineage reveals no corresponding periodic repeat. The three types of cell are generated semiclonally (Fig. 13) and the repeat units are formed by reproducible but piecemeal recruitment. Presumably the regularity of the structure depends upon cell affinity and perhaps upon packing constraints in the embryo. There appears to be no correlation between the repetition in the hypodermis and that in the endoderm, muscle, or juvenile nervous system. During the Postembryonic expansion of the ventral nervous system (Sulston and Horvitz, 1977), the regularly repeated array of neurons derived from the P cells is simply superimposed upon the different, and less regular, repeat pattern of juvenile neurons.
This mode of limited "segmentation" partially parallels the true segmentation of the leech, which also develops by an invariant cell lineage (Stent and Weisblat, 1982). The leech embryo has five stem cells (four ectodermal and one mesodermal) on either side; every segment is founded by a group of cells comprising one or two daughters from each of the stem cells. Consequently, as in the nematode, a given cell in the mature animal is more closely related to its homologues in other segments than it is to most of the cells in its own segment.
Sublineages
We shall use the term "sublineage" as an abbreviation for the more descriptive, but cumbersome, phrase "intrinsically determined sublineage" - namely, a fragment of the lineage which is thought to be generated by a programme within its precursor cell. At present there is no direct evidence for such inherent programming, and so strictly the concept is hypothetical; nevertheless, in certain cases a variety of circumstantial evidence can be adduced to support it. The notion has been discussed elsewhere (Chalfie et al, 1981; Sternberg and Horvitz, 1982) and need not be further elaborated now, but it is convenient to begin by listing two of the available criteria for postulating the existence of a sub-lineage: (1) the generation of the same lineage, giving rise to the same set of cell fates, from a series of precursors of diverse origin and position; (2) evidence for cell autonomy within the lineage, obtained from laser ablation experiments or the study of mutants.
By the first criterion, two sublineages can be found in the head of the embryo (Fig. 19). The sixfold inner labial sublineage generates the neurons of the six identical inner labial sensilla. The outer labial sublineage can be regarded as either sixfold or (four + two)fold; this ambiguity is of interest in the light of the morphological difference between the OLQ and OLL outer labials (Fig. 19 legend; White et al, in preparation).
 Figure 19. Diagrammatic longitudinal section through spicule of a late L4 male (42 hr, series 1). Tracings of electron micrographs of transverse sections at three levels are included to show the detailed arrangement of the cells. The two ciliated neurons (SPD and SPV) open to the exterior. The two sheaths (SPshD and SPshV) have already begun to fuse at their tips.
Figure 19. Diagrammatic longitudinal section through spicule of a late L4 male (42 hr, series 1). Tracings of electron micrographs of transverse sections at three levels are included to show the detailed arrangement of the cells. The two ciliated neurons (SPD and SPV) open to the exterior. The two sheaths (SPshD and SPshV) have already begun to fuse at their tips.
In postembryonic development, parts of the lineage which are simply bilaterally symmetrical do not usually qualify as sublineages by the first criterion above, because their precursors are of symmetrical origin. However, in the case of the embryo it is possible to take the analysis a step further.
In the lineage chart (Fig. 3), pairs of precursors which give rise to similar or identical division patterns and groups of cells on the left and right sides are indicated. In the posterior part of the animal, bilateral symmetry arises at an early stage, as a result of equational divisions of precursors, and the groups are large. In the anterior part, however, symmetry arises late, in a piecemeal fashion, and the groups are small; for example, identical lineages arise from ABalaappp and ABalapaap, which are not themselves of identical origin. There are therefore grounds for supposing that these symmetrical pieces of lineage are Sublineages, particularly since their precursors are frequently not disposed symmetrically and do not have identical neighbours on the left and the right.
The latter observations suggest further that the precursors are determined, not by interaction with their neighbours, but by their ancestry. Evidence in favour of this conjecture was obtained from a series of laser ablation experiments, in which all tested precursors behaved autonomously. Some implications of this finding are discussed below (Symmetry and Asymmetry).
The assignment of sublineages is often a matter for subjective judgment, in that two sections of the lineage may be alike in some respects and discrepant in others. Thus, ABalpaaa and ABarapaa give rise to sublineages identical except for the fate of one cell, and it is reasonable to suppose that identical programmes are being used by the two precursors; the fate of the one anomalous cell may be determined either by position or by additional intrinsic factors. A more extreme example is provided by the pair ABalapap and ABalappp; in this case a dividing cell in one version of the sublineage is replaced by a cell death in the other, but there are still enough common features to suggest a shared programme.
Programmed Cell Death and Sexual Dimorphism
The large number of programmed cell deaths, and their reproducibility, is evident from the lineage. The most likely reason for the occurrence of most of them is that, because of the existence of sublineages, unneeded cells are frequently generated along with needed ones (Sulston and Horvitz, 1977; Horvitz et al, 1982).
Direct support for this point of view is provided by the sexually specific deaths seen in the embryonic lineage. In the hermaphrodite the putative CEM cells die but the HSN cells survive, whilst in the male the situation is reversed; before the deaths, however, each cell begins to differentiate in the same way regardless of sex. Rather than generate two different patterns of cell division, the nematode uses the same sublineages in both sexes and kills those cells that are not required. Sexually specific deaths are also seen in hermaphrodite larvae, but in other cases no surviving homologues of the dead cells are available for study. However, by means of mutants in which cell death does not occur (recently isolated by H. Ellis, personal communication; Horvitz et al., 1982), it should now be possible to exhume all of them.
There are few obvious restrictions upon the patterns of cell deaths. They are found in ectoderm, mesoderm, and (in Turbatrix and Panagrellus) endoderm. Several precursors give rise to two cell deaths in successive divisions, but none give rise to two in a single division. Presumably there is some selective pressure on the organism to dispense with programmed deaths, which must represent a waste of time and materials. Logically, sister deaths could be readily eliminated (by programming the death of their parent) whereas single deaths might require more extensive rearrangement of control elements. Sternberg and Horvitz (1981) have reported sister deaths in the Panagrellus gonad, but suggest, on account of their variability, that they are of recent evolutionary origin.
Rotational Symmetry
In addition to bilateral symmetry (see Sublineages) parts of the nematode display two-, three-, four-, and sixfold rotational axes of symmetry. The embryonic lineage, on the other hand, is in part bilaterally symmetrical and in part asymmetrical; the ways in which it generates the more elaborate symmetries of the mature nematode will now be discussed.
Twofold. At 300 min the gonad is a bilaterally symmetrical structure consisting of two identical subunits, each comprising two cells of different types (Fig. 16). It subsequently turns to lie obliquely and acquires a twofold rotational axis of symmetry. This behaviour can be understood if, during late embryogenesis, the two subunits interact specifically with one another but not with nongonadal tissues, the oblique position being a consequence of packing. The autonomy of the gonad was pointed out by Kimble and Hirsh (1979) who found that, apart from the initial events in the male, post-embryonic gonadogenesis is reproducibly oriented with regard to the axes of the gonad rather than to those of the body.
Threefold. The muscles and structural cells of the pharynx have a precise threefold rotational axis of symmetry, whilst the nervous system of the pharynx is bilaterally symmetrical. Broadly speaking, two of the three identical sets of mechanical elements are generated by bilaterally symmetrical lineages, and the third set is assembled by piecemeal recruitment. The threefold symmetrical arrays of the arcade, hypl, hyp4, and parts of the valves are produced in a similar fashion. This plan of development contrasts with that found for the vas deferens (formed in the larval male), in which structures having a threefold rotational axis of symmetry are generated by three equivalent precursor cells which give rise to identical sublineages (Kimble and Hirsh, 1979; Kimble, 1981).
Four- and sixfold. At the tip of the head the various classes of sensilla are arranged in rings with bilateral (amphid), fourfold (cephalic), and sixfold symmetry (inner labial); the outer labial sensilla can be regarded as having either sixfold or (four + two)fold symmetry. Various authors have speculated that one of the higher modes of symmetry may represent the fundamental plan of the nematode head (discussed by Ward et al, 1975). If this were the case, the plan should be detectable as a corresponding pattern of precursor cells. However, apart from the sublineages giving rise to the labial neurons (see Sublineages), no mode of symmetry other than bilateral is detectable in the embryonic lineage. Even the origin of the labial neurons does not support the idea that sixfold symmetry is fundamental, because the sublineages for the inner and outer labial sensilla are generated by unrelated precursors and do not include the supporting cells.
Symmetry and Asymmetry
It is clear from the events already discussed that, at numerous points in the lineage, symmetry is broken. This is obviously a necessary step on the pathways to unique cells and threefold symmetrical structures, but it can also occur in the generation of bilaterally symmetrical structures (see Sublineages).
More than one symmetry change can occur during the ancestry of a single cell. An extreme example is provided by a sequence from the complete (embryonic and Postembryonic) lineage of the male; in it, symmetry is generated three times and broken twice, as follows. Cells B and U are produced by identical right and left lineages; they break symmetry and arrange themselves respectively posterior and anterior to the anus; during larval development, the anterior daughter of B divides symmetrically and gives rise to identical lineages on the left and right sides; two pairs of its progeny break symmetry again and arrange themselves anterior-posteriorly; finally, all four of these progeny generate symmetrical groups of cells on the two sides of the animal (Sulston et al, 1980).
In addition to symmetry changes involving precursors, which are indicated in Fig. 3, certain new symmetries appear at the level of terminal differentiation (Fig. 20). These "phase shifts" in otherwise symmetrical patterns of fate assignments are reminiscent of those seen in the postembryonic development of male-specific muscle (Sulston et al, 1980; Sulston and White, 1980). They may have arisen as a result of selective pressure to produce either unique cells or threefold groups, and various hypothetical schemes can be constructed to explain their evolution. Sternberg and Horvitz (1981) have termed such events "altered segregations," to express the observation that two cells may acquire the same developmental potential by different route
 Figure 20. Neurons of Dx class passing through basement membranes(arrowheads) and forming electrical junctions with muscle cells (mu). Electron micrograph of transverse section series 4. Bar = 1 µm.
Figure 20. Neurons of Dx class passing through basement membranes(arrowheads) and forming electrical junctions with muscle cells (mu). Electron micrograph of transverse section series 4. Bar = 1 µm.
How are the symmetry decisions taken? Laser ablation experiments show that two cases of symmetry breakage (G2/W and duct/G1, see Cell Interaction Experiments) involve competitive interaction between the homologous partners, which confront one another at the midline. In all other cases tested the behaviour of the cells seems to be independent of their neighbours and of their exact position. We are therefore forced to conclude that the various symmetries arise during development largely from a series of piecemeal decisions, each individually selected in the course of evolution. The precise bilateral symmetry of the head, for example, is not determined by a single global influence (such as a symmetrical morphogenetic field) but is the end result of many small decisions. Bilateral symmetry is evidently a favourable asset for an animal, and is selected even at the level of individual neurons and sensilla.
Conclusion
Perhaps the most striking findings are firstly the complexity and secondly the cell autonomy of the lineage. As regards the former, many of the guesses that have been made in the past about the ultimate fates of embryonic cells are correct in general but wrong in detail: the assignment of cell function follows certain broad rules to which there are numerous exceptions. As regards the latter, there are two examples of specific interaction between cells affecting their fates, and there is evidence for induction late in embryogenesis; doubtless other instances of cell-to-cell signalling remain to be discovered, but much of the detailed control of division pattern and fate, including such niceties as the generation of precise bilateral symmetry, seems to be vested in autonomous programmes within individual cells.
With hindsight, we can rationalise both this complexity and this rigidity. The nematode belongs to an ancient phylum, and its cell lineage is a piece of frozen evolution. In the course of time, new cell types were generated from precursors selected not so much for their intrinsic properties as for the accident of their position in the embryo. Perhaps the mutations responsible for these novel developmental events acted directly upon the intrinsic behaviour of the precursors, but equally they may have operated via extrinsic regulative mechanisms. In the latter case, however, cell-cell interactions that were initially necessary for developmental decisions may have been gradually supplanted by autonomous programmes that were fast, economical, and reliable, the loss of flexibility being outweighed by the gain in efficiency. On this view, the perverse assignments, the cell deaths, the long-range migrations -all the features which could, it seems, be eliminated from a more efficient design- are so many developmental fossils. These are the places to look for clues both to the course of evolution and to the mechanisms by which the lineage is controlled today.
Appendix
PARTS LIST: Caenorhabditis elegans (Bristol), NEWLY HATCHED LARVA
This index was prepared by condensing a list of all cells in the adult animal, then adding comments and references. A complete listing is available on request to MRC, Cambridge. References: (1) Ward et al, 1975; (2) Ware et al, 1975; (3) White et al, 1976; (4) Albertson and Thomson, 1976; (5) Singh and Sulston, 1978; (6) Kimble and Hirsh, 1979; (7) Sulston et al, 1975; (8) Chalfie and Sulston, 1981; (9) Sulston and Horvitz, 1977; (10) Sulston et al, 1980; (11) White et al, 1978; (12) Horvitz et al, 1982; (13) Wright and Thomson, 1981; (14) Gossett et al, 1982; (15) Nelson et al, 1983.
PARTS LIST PAGE 1
PARTS LIST PAGE 2
PARTS LIST PAGE 3
PARTS LIST PAGE 4
PARTS LIST PAGE 5
PARTS LIST PAGE 6
PARTS LIST PAGE 7
PARTS LIST PAGE 8
PARTS LIST PAGE 9
Acknowledgments
We are greatly indebted to numerous colleagues for discussions, for sharing their results, and for commenting on the manuscript. In particular, we wish to acknowledge Gunter von Ehrenstein, who died on 26 December 1980, and in whose laboratory part of this work was carried out; Sydney Brenner, who is uniquely responsible for generating the current interest in the development of C. elegans; Eileen Southgate, for help with the interpretation of electron micrographs; Donna Albertson, Randall Cassada, Martin Chalfie, Antonio Garcia-Bellido, Ed Hedgecock, Jonathan Hodgkin, Bob Horvitz, Cynthia Kenyon, Judith Kimble, Peter Lawrence, Paul Sternberg, and Michael Wilcox for advice and comments.
References
Albertson, D. G., and Thomson, J. N. (1976). The pharynx of Caenorhabditis elegans. Phil. Trans. Roy. Soc. London Ser. B 275, 299-325.
Albertson, D. G., and Thomson, J. N. (1982). The kinetochores of Caenorhabditis elegans. Chromosoma 86, 409-428.
Toveri, T. (1892). Uber die Entstehung des Gegensatzes zwischen den Geschlechtszellen und den somatischen Zellen bei Ascaris megalocephala. Sitzungsber. Ges. MorphoL PhysioL 8, 114-125.
Brenner, S. (1974). The genetics of Caenorhabditis elegans. Genetics 77, 71-94.
Chalfie, M., Horvitz, H. R., and Sulston, J. E. (1981). Mutations that lead to reiterations in the cell lineages of C. elegans. Cell 24, 59-69.
Chalfie, M., and Sulston, J. (1981). Developmental genetics of the mechanosensory neurons of Caenorhabditis elegans. Develop. Biol. 82, 358-370.
Chitwood, B. G., and Chitwood, M. B. (1974). "Introduction to Nematology." Univ. Park Press, Baltimore.
Deppe, U., Schierenberg, E., Cole, T., Krieg, C., Schmitt, D., Yoder, B., and von Ehrenstein, G. (1978). Cell lineages of the embryo of the nematode Caenorhabditis elegans. Proc. Nat. Acad. Sci. USA 75, 376-380.
Garcia-bellido, A., Ripoll, P., and Morata, G. (1973). Developmental compartmentalisation of the wing disk of Drosophila. Nature New Biol. 245, 251-253.
Gossett, L. A., Hecht, R. M., and Epstein, H. F. (1982). Muscle differentiation in normal and cleavage-arrested mutant embryos of Caenorhabditis elegans. Cell 30, 193-204.
Hirsh, D., Oppenheim, D., and Klass, M. (1976). Development of the reproductive system of Caenorhabditis elegans. Develop. Biol. 49, 200-219.
Horvitz, H. R., Chalfie, M., Trent, C., Sulston, J. E., and Evans, P. D. (1982). Serotonin and octopamine in the nematode Caenorhabditis elegans. Science 216, 1012-1014.
Horvitz, H. R., Ellis, H. M., and Sternberg, P. W. (1982). Programmed cell death in nematode development. Neurosci. Comment. 1, 56-65.
Kimble, J. (1981). Alterations in cell lineage following laser ablation of cells in the somatic gonad of Caenorhabditis elegans. Develop. Biol. 87, 286-300.
Kimble, J., and Hirsh, D. (1979). The Postembryonic cell lineages of the hermaphrodite and male gonads in Caenorhabditis elegans. Develop. Biol. 70, 396-417.
Kimble, J. E., and White, J. G. (1981). On the control of germ cell development in Caenorhabditis elegans. Develop. Biol 81, 208-219.
Klass, M., Wolf, N., and Hirsh, D. (1976). Development of the male reproductive system and sexual transformation in the nematode Caenorhabditis elegans. Develop. Biol 52, 1-18.
Krieg, C., Cole, T., Deppe, U., Schierenberg, E., Schmitt, D., Yoder, B., and von Ehrenstein, G. (1978). The cellular anatomy of embryos of the nematode Caenorhabditis elegans. Develop. Biol 65, 193-215.
Laufer, J. S., Bazzicalupo, P., and Wood, W. B. (1980). Segregation of developmental potential in early embryos of Caenorhabditis elegans. Cell 19, 569-577.
Laufer, J. S., and von Ehrenstein, G. (1981). Nematode development after removal of egg cytoplasm: Absence of localised unbound determinants. Science 211, 402-405.
Lawrence, P. A. (1981). The cellular basis of segmentation in insects. Cell 26, 3-10.
Le Douarin, N. (1980). Migration and differentiation of neural crest cells. In "Current Topics in Developmental Biology" (A. A. Moscona and A. Monroy, eds.), Vol. 16, pp. 31-85. Academic Press, New York.
Mounier, N. (1981). Location of neurosecretory-like material in Caenorhabditis elegans. Nematologica 27, 160-166.
Nelson, F. K., Albert, P. S., and Riddle, D. L. (1983). Fine structure of the Caenorhabditis elegans secretory-excretory system. J. Ultrastruct Res. 82, 156-171.
Nigon, V. (1965). Developpement et Reproduction des Nematodes. In "Traite de Zoologie" (P.-P. Grasse, ed), Vol. 4, pp. 218-294.
Pai, S. (1927). Lebenzyklus der Anguillula aceti. Ehrbg. ZooL Anz. 74, 257-270.
Ritter, M. (1965). in "Traite de Zoologie" (P.-P. Grasse, ed.), Vol. 4, p. 782. Masson & Cie, Paris.
Robertson, A. M. G., and Thomson, J. N. (1982). Morphology of programmed cell death in the ventral nerve cord of Caenorhabditis elegans larvae. J. Embryol. Exp. Morphol 67, 89-100.
Seck, P. (1938). Zur Entwicklungsmechanik des Essigalchens. Arch. Entwicklungsmech. 137, 57-85.
Schierenberg, E., and Cassada, R. (1982). Cell division patterns and cell diversification in the nematode Caenorhabditis elegans. In "Identification and Characterisation of Stem Cell Populations" (C. S. Potten, ed.). Churchill Livingstone, Edinburgh.
Singh, R. N., and Sulston, J. E. (1978). Some observations on moulting in Caenorhabditis elegans. Nematologica 24, 63-71.
Stent, G. S., and Weisblat, D. A. (1982). The development of a simple nervous system. Sci. Amer. 246(1), 100-110.
Sternberg, P. W., and Horvitz, H. R. (1981). Gonadal cell lineages of the nematode Panagrellus redivivus and implications for evolution by the modification of cell lineage. Develop. Biol 88,147-166.
Sternberg, P. W., and Horvitz, H. R. (1982). Postembryonic non-gonadal cell lineages of the nematode Panagrellus redivivus: Description and comparison with those of Caenorhabditis elegans. Develop. Biol 93, 181-205.
Strome, S., and Wood, W. B. (1982). Immunofluorescence, visualization of germ-line-specific cytoplasmic granules in embryos, larvae, and adults of Caenorhabditis elegans. Proc. Nat. Acad. Sci. USA 79, 1558-1562.
Sulston, J. E., Albertson, D. G., and Thomson, J. N. (1980). The Caenorhabditis elegans male: Postembryonic development of non-gonadal structures. Develop. Biol 78, 542-576.
Sulston, J., Dew, M., and Brenner, S. (1975). Dopaminergic neurons in the nematode Caenorhabditis elegans. J. Comp. Neurol 163, 215-226.
Sulston, J. E. and Horvitz, H. R. (1977). Post-embryonic cell lineages of the nematode Caenorhabditis elegans. Develop. Biol 56, 110-156.
Sulston, J. E., and Horvitz, H. R. (1981). Abnormal cell lineages in mutants of the nematode Caenorhabditis elegans. Develop. Biol 82, 41-55.
Sulston, J. E., and White, J. G. (1980). Regulation and cell autonomy during Postembryonic development of Caenorhabditis elegans. Develop. Biol 78, 577-597.
von Ehrenstein G., Sulston, J. E., Schierenberg, E., Laufer, J. S., and Cole, T. (1981). Embryonic cell lineages and segregation of developmental potential in Caenorhabditis elegans. In "International Cell Biology 1980-1981." Springer-Verlag, Berlin/New York.
Ward, S., Thomson, N., White, J. G., and Brenner, S. (1975). Electron microscopical reconstruction of the anterior sensory anatomy of the nematode Caenorhabditis elegans. J. Comp. Neurol 160, 313-338.
Ware, R. W., Clark, C., Crossland, K., and Russell, R. L. (1975). The nerve ring of the nematode Caenorhabditis elegans. J. Comp. Neurol 162, 71-110.
White, J. G. (1974). "Computer-Aided Reconstruction of the Nervous System of C. elegans." Ph.D. thesis, University of Cambridge.
White, J. G., Albertson, D. G., and Anness, M. A. R. (1978). Connectivity changes in a class of motoneurone during the development of a nematode. Nature (London) 271, 764-766.
White, J. G., Southgate, E., Thomson, J. N., and Brenner, S. (1976). The structure of the ventral nerve cord of Caenorhabditis elegans. Phil Trans. Roy. Soc. London Ser. B 275, 327-348.
Wright, K. A., and Thomson, J. N. (1981). The buccal capsule of Caenorhabditis elegans (Nematoda: Rhabditoidea): An ultrastructural study. Canad. J. ZooL 59, 1952-1961.
zur Strassen, 0. (1892). Bradynema rigidum v. Sieb. Z. Wissenschaften. ZooL 54, 655-674.
zur Strassen, 0. (1959). Neue Beitrage zur Entwicklungsmechanik der Nematoden. Zoologica 107, 1-142.
|