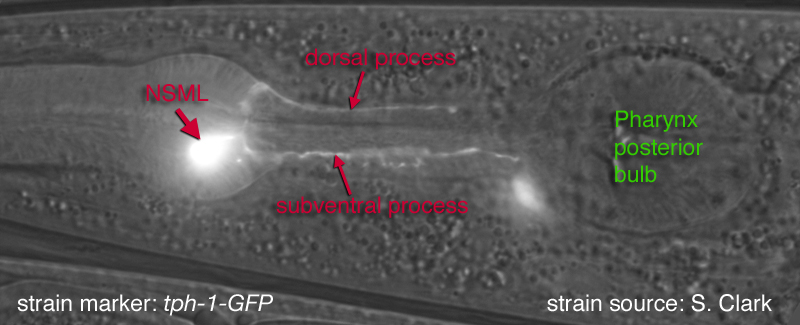
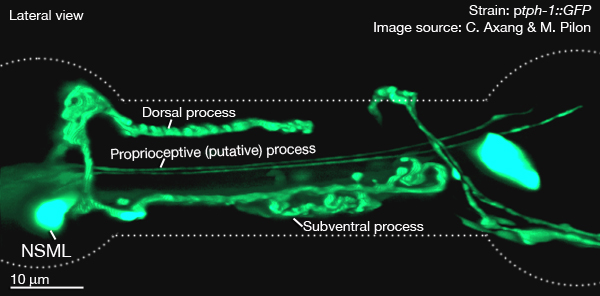
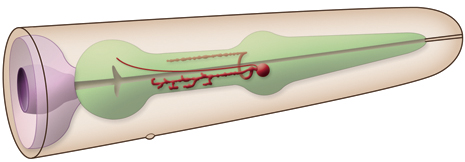
of the isthmus. The other branch runs posteriorly within the subventral nerve cord of the isthmus.These processes run at the most outside edge of the nerve cords, very close to pseudocoelom, and make synapses to the pharyngeal basement membrane and muscle cells. Both processes terminate before reaching the terminal bulb. Within the isthmus these processes periodically swell to form varicosities filled with large, lightly- and darkly-staining membrane bound vesicles, as well as small clear vesicles.
Along with these major processes NSM's also have small, synapse-free processes with free endings just under the cuticle of the pharyngeal lumen at the boundary between the corpus and isthmus (where bacteria accumulate). These endings are attached to the adjacent muscle cells by desmosomes and are thought to act as stretch (proprioceptive) receptors. These processes can not be seen at the light microscopic level (therefore not obvious in GFP images below). They have also been omitted from the drawn images below. The NSM's receive extensive synapses from I2 (see connectivity diagram below) (Albertson and Thomson, 1976). NSM's are serotonergic but they are also able to take up exogenous serotonin via a serotonin reuptake transporter, MOD-5 (Ranganathan et al., 2001)
Neurotransmitter/ Neuropeptide:
- Serotonin
- NLP-13; neuropeptide-like protein
- NLP-18; neuropeptide-like protein
- NLP-19; neuropeptide-like protein
[N.B. According to recent studies, glutamate does not seem to be a neurotransmitter in NSMs, although some weak and variable EAT-4 (VGluT) expression was noted in a previous work (Lee et al., 1999; O. Hobert, pers. comm.)]
(Nathoo et al., 2001; Horvitz et al., 1982; Sze et al., 2000)
Innexin expression:
- UNC-9
(Altun et al., 2009)
Receptor expression:
- GLR-7; glutamate receptor subunit
- GLR-8; glutamate receptor subunit
- SER-2; a splice variant of the tyramine receptor
- Possibly AEX-2; G protein-coupled receptor
(Tsalik et al., 2003; Round et al., 2002; Brockie et al., 2001)
Function:
- These neurons were suggested to have neurohumoral function signaling the presence of food to the body since NSM's appear secretory by morphology and have processes that run in close apposition to the pseudocoelom over most of their lengths (Albertson and Thomson, 1976, Avery and Thomas, 1997).
- They were postulated to sense bacteria in the pharynx lumen by their proprioceptive endings and transmit this (presence of food) to the rest of the body by secretions to the pseudocoelomic fluid. The presence of food works as a signal for increased pharyngeal pumping, decreased locomotion and increased egg-laying behavior. Exogenous application of serotonin also induces these responses supporting the hypothesis NSM's can function neurohumorally by serotonin secretion. However, ablation of NSM's have only subtle effects on pumping suggesting they may not play any important role in this behavior (Avery et al., 1993, Avery and Thomas, 1997).
- May modulate "the enhanced slowing response"; when placed on a bacterial lawn, previously food-deprived animals slow down their locomotion to a higher degree compared to well-fed animals. This phenomenon is described as "the enhanced slowing response". When NSM's are ablated there is a small but significant decrease in the enhanced slowing response suggesting NSM's contribute to this behavior (Sawin et al., 2000).
|




 Click pictures for higher resolution images
Click pictures for higher resolution images