Neurotransmitter/ Neuropeptide:
- Glutamate (AWC-AIA synapse is an inhibitory glutamatergic synapse whose function is acutely modulated by NLP-1/NPR-11 signaling)
- NLP-1; neuropeptide-like protein
(Chalasani et al., 2010; Serrano-Saiz et al., 2013)
Innexin expression:
- CHE-7
- INX-1a
- INX-1b
- INX-7
- INX-10a
- INX-14
- INX-18a
- INX-19
- UNC-7
- UNC-9
(Chuang et al, 2007; Bhattacharya et al., 2019)
Receptor expression:
- OSM-9; mammalian capsaicin receptor-like channel protein
- DAF-11; transmembrane guanylyl cyclase
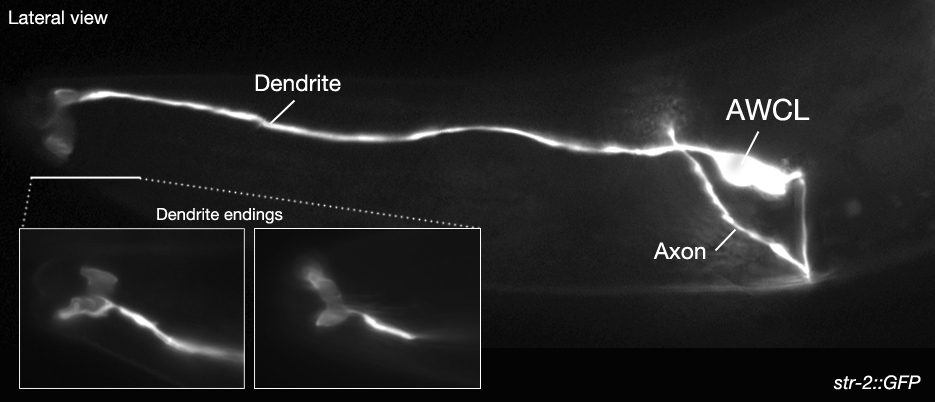
- STR-2, G protein-coupled serpentine receptor, expressed asymmetrically (randomly on the right or left side)
(Birnby et al., 2000; Troemel et al., 1999; Colbert et al., 1997)
Function:
- Critical for chemotaxis to volatile odorants, (e.g. chemotaxis to benzaldehyde, butanone, isoamylalcohol, 2,3 pentanedione and 2,4,5 trimethylthiazole).
- Induction of local search behavior and promoting turns. After animals are removed from bacterial food, they initiate a local search behavior consisting of reversals and deep omega-shaped turns. This is followed by dispersal ~30 min later as reversal and turns are suppressed. Local search behavior is triggered by AWC olfactory neurons, ASK gustatory neurons, and AIB interneurons, while dispersal is promoted by ASI gustatory neurons and AIY interneurons (Gray et al., 2005). The AWC neurons synapse onto several interneurons including AIB and AIY, which enhance and suppress turning, respectively. The AWC neurons activate the AIB interneurons through AMPA-type glutamate receptors and inhibit AIY interneurons through glutamate-gated chloride channels. AWC are odor-OFF neurons that are activated by odor removal and inhibited in the continued presence of odors. On prolonged exposure to odorant (over 30 min), adaptation to AWC-sensed odors occurs, abrogating the chemotactic response. NLP-1 is required for these AWC-dependent food-evoked behaviors and odor adaptation (Chalasani et al., 2010; Chalasani et al., 2007; Gray et al., 2005; L'Etoile and Bargmann, 2000; Bargmann et al., 1993).
- Thermosensor: After cultivation at a uniform temperature (Tc) with sufficient food, animals preferentially migrate to their cultivation temperature (Tc) when placed on a thermal gradient, and move isothermally at this temperature (Hedgecock and Russell 1975), animals sense and record their Tc by AFD (major thermosensory) and AWC and ASI neurons (Beverly et al, 2011; Kuhara et al., 2008; Biron et al., 2008). This Tc memory is plastic and can be reset upon cultivation at a different temperature (Hedgecock and Russell 1975). Activity of both the AFD and the AWC neurons is essential for the execution of thermotactic behaviors with high fidelity and precision. Upon sensation of a higher temperature than Tc, Ca++ concentration in AWC increases via production of cGMP by ODR-1 and activation of cGMP-dependent TAX-4 cation channel (Kuhara et al., 2008). Unlike AFD neurons, which respond to thermal stimuli above Tc with continuous, graded calcium signals in a deterministic and highly reproducible way, AWC neurons exhibit stochastic temperature-evoked Ca++ changes that are stimulus-correlated with different temporal kinetics at temperatures above and below Tc (Biron et al., 2008). AWC appears to get more active as temperatures rise or fall further from Tc. The AFD, AWC and ASI neurons seem to act in concert to increase turning rate when animals encounter higher temperatures than Tc on a gradient to move down the gradient toward colder temperatures near Tc (negative thermotaxis) (Biron et al., 2008). To track isotherms, animals do not actively pursue isothermal alignment, but once serendipitously aligned along an isotherm (at T=Tc), they track by suppressing turns (Luo et al., 2006). G-protein-coupled receptor (GPCRs) SRTX-1 is required both in AFD and AWC for retaining normal isothermal tracking (Liu et al., 2012). AWC neurons are relatively quiet around temperatures close to the Tc.
- Electrosensory navigation; C. elegans moves toward the negative pole of an electric field. Killing the ASJ or ASH neurons leads to significant disruption in electrotaxis, while killing ASK, AWB or AWC has a weaker effect (Gabel et al., 2007).
- One of the three core sensory neurons (AWA, AWC, ASK) that are required for sexual-attraction in males (White and Jorgensen, 2012).
Reporters:
- At CGC: OH16052 otIs745 [ceh-36p3::npp-9::mCherry::BLRP::3xFLAG::npp-9 3'UTR]
- Other:
|





 Click pictures for higher resolution images
Click pictures for higher resolution images