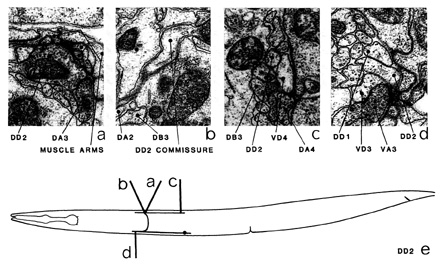
DDn is a set of six motoneurons, with cell bodies in the ventral cord, which innervate dorsal
muscles. Each cell has a short, posteriorly directed and a longer, anteriorly directed process
emanating from its cell body. The anterior process has a branch, which leaves the ventral cord
as a commissure and runs round to the dorsal cord. (The DD1 commissure runs to the left,
the rest to the right.) The commissures often run through the hypodermal ridge when entering
or leaving the nerve cords (b). The commissure splits, as it enters the dorsal cord, into an
anteriorly and a posteriorly directed process, which span approximately the same longitudinal
region of the body as their ventral counterparts. Both the dorsal and the ventral processes run
adjacent to the basal lamina bounding the cord and run in close association with the processes
of the VDn neurons. In the ventral cord they run between the more dorsal processes of the
VC neurons and the more ventral processes of the VAn and VBn neurons (figure 18). In the
dorsal cord they run to ventral of the other motoneuron classes (figure 19). The DDn processes
in the ventral cord are exclusively postsynaptic and receive synaptic input from VAn (d), VBn (*a) and VCn (*b) as corecipients at NMJs, often with short dendritic spines intercepting the
NMJs (as in VAn-a). The processes in the dorsal cord are predominantly presynaptic and have
many NMJs (a), most of which have only muscle as the postsynaptic partner. The role of the
dorsal and ventral processes is reversed in the first larval stage and switches round to the adult
configuration at about the time of the first moult (White et al. 1978). The processes of DDn and VDn do not have extended, apparently undifferentiated, distal regions, as do the other
classes of motoneuron; instead, they end abruptly in close proximity to the end of a
neighbouring process of the same class and there is usually a gap junction between them (d). There are also a few gap junctions to VDn neurons (c).
Magnifications: (a-d) x 25500.


Click pictures for higher resolution images