|
MUSCLE SYSTEM OF THE MALE - OVERVIEW
 Click pictures for new window with figure and legend Click pictures for new window with figure and legend
1 General Information
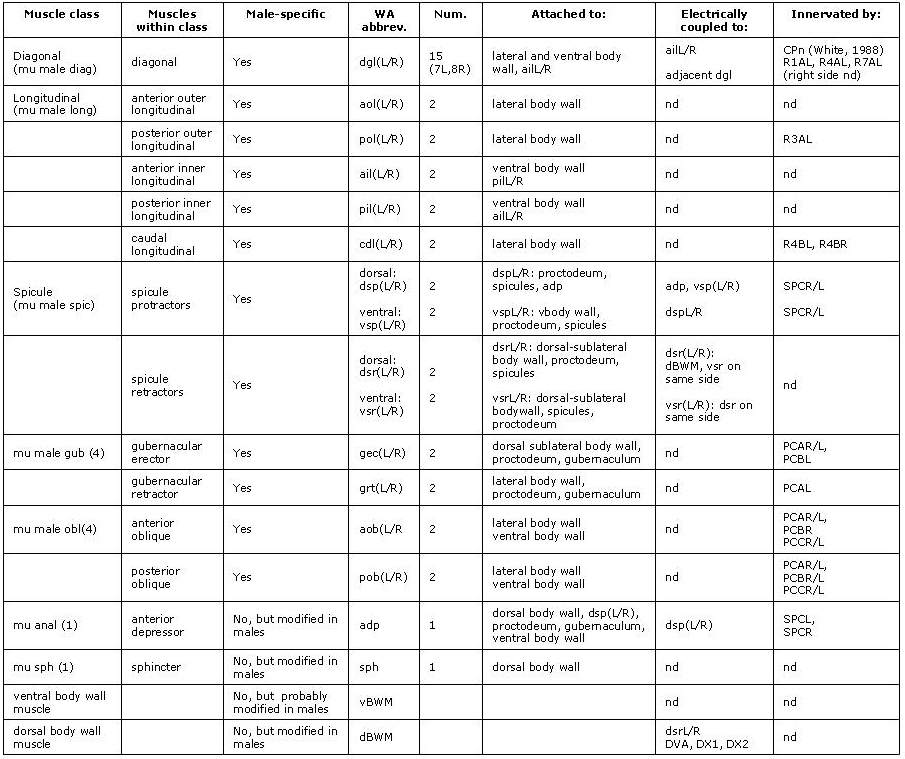
The muscle system of the male differs from that of the hermaphrodite primarily in its complement of sex muscles, derived from the M lineage. The adult male has 41 M-derived sex muscles consisting of 7 morphologically distinct types. None of these muscles are present in the hermaphrodite and so they are male-specific muscles. These muscles are distributed through the posterior half of the animal and function in various aspects of male mating behavior or copulation (MaleMusFIG 1, MaleMusFIG 2 and MaleMusMOVIE 1; Sulston and Horvitz, 1977; Sulston et al., 1980). Indeed, several sex muscles are directly associated with structures and tissues of the male copulatory apparatus such as the spicules and the gubernaculum (see also Male-specific muscles). Surprisingly, males in which M has been ablated can execute most of the initial steps of mating (backing, turning, vulval location; Sulston, J.E., WBG 7(2):22), although their execution is significantly poorer than in wild type (see Loer and Kenyon, 1993). The spicules in such ablated animals do not function. Thus some male-specific muscles are essential for mating (e.g. the spicule muscles) while others appear to enhance the animal's ability adopt certain postures required during mating.
 In addition to the sex muscles, a few embryonically-derived muscles that are present in both males and hermaphrodites (sphincter, anal depressor and body wall muscles) express different characteristics in the male and are employed in mating behavior or copulation (MaleMusFIG 1B, underlined labels and MaleMusMOVIE 1; Sulston and Horvitz, 1977; Sulston et al., 1980; Sulston et al., 1983).
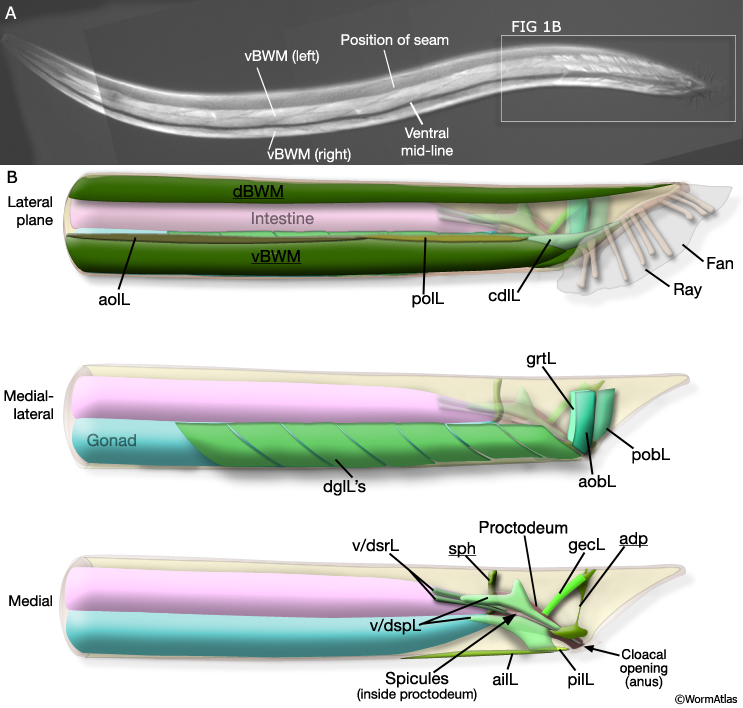
MaleMusFIG 1: The adult male sex muscles. A. Epifluorescent images from transgenic animal expressing the unc-27 ::GFP reporter gene (Troponinl), left lateral oblique view. (Strain source: L. Jia and S.W. Emmons.) B. Illustrations highlighting area boxed in A, left lateral view. Sexually dimorphic areas are underlined. Lateral (dBWM and vBWM) Dorsal and ventral body wall muscle; (aolL and polL) anterior and posterior longitudinal, left; (cdlL) caudal outer longitudinal, left. Medial-lateral (dglL) Diagonal muscles, left; (grtL) gubernacular retractor, left; (aobL and pobL) anterior and posterior oblique, left. Medial (v/dsrL) Ventral and dorsal spicule retractors, left; (v/dspL) ventral and dorsal spicule protractors; (sph) sphincter; (ailL and pilL) anterior and posterior inner longitudinal; (gecL) gubernacular erector, left; (adp) anal depressor.
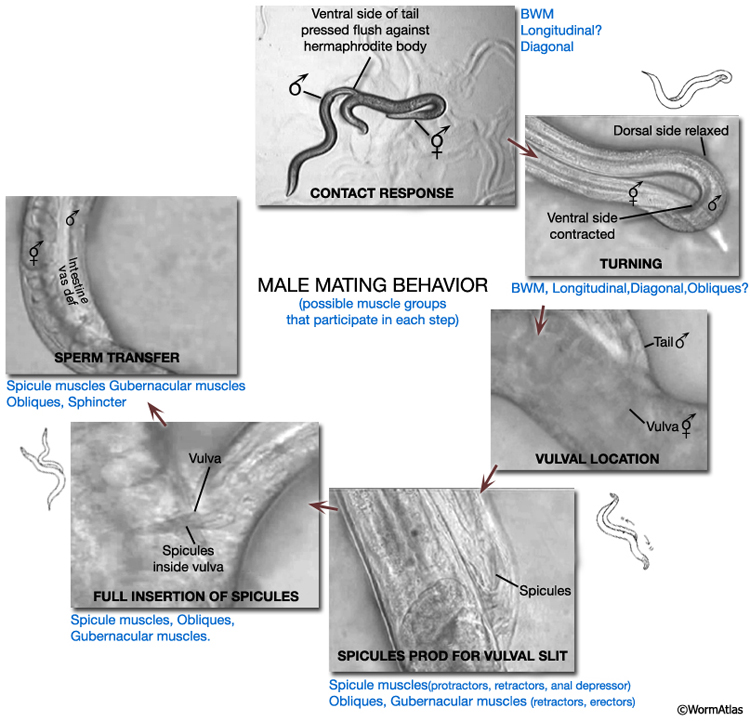
MaleMusFIG 2: Male mating behavior. Steps in mating featuring participating muscle groups. (Based on Loer and Kenyon, 1993; Liu and Sternberg, 1995; Barr and Sternberg, 1999; Garcia et al., 2001.)
With the exception of the longitudinal muscles, the (M-derived) sex muscles of the male generally conform to the non-striated class of muscle types (see Muscle System - Introduction for a description of muscle types and Muscle System - Nonstriated Muscle for hermaphrodite nonstriated muscles. In contrast to striated muscles (e.g. body wall muscles), non-striated muscles contain a single or only a few sarcomeres (contractile units) and attach to cuticle only at their ends (for organization of attachment sites see CutFIG 10, The Cuticle; MusFIG 11 and Somatic Muscle). The cuticle acts as an exoskeleton for muscle contraction to work against. In the male the cuticle may be body wall cuticle, as in the hermaphrodite, or a male-specific cuticle such as the spicules or the gubernaculum (MaleMusTABLE 1). Certain subsets of muscles in the male are also connected to each other via attachment plaques (MaleMusTABLE 1; denoted by yellow arrows in chapter figures). These muscle::muscle connections share some molecules in common with cuticle::muscle connections (see Muscle System - Somatic Muscle). While the bulk of contractile force is likely to be transmitted to cuticle, muscle::muscle connections may transmit some pulling force between interconnected muscles, enhancing their ability to function as a unit (for example the spicule protractors and anal depressor muscles). Also like muscles elsewhere in the body, the male sex muscles have muscle arms and these are often the sites for innervation. However, the arms can be short and in some cases synapses are made in the absence of arms, directly on to the muscle sarcomere region (e.g. spicule protractors, Male-specific Muscles).
2 Development of the Male Muscle System
Sexual specialization of the male muscle system beings in L2 and continues through to the final (L4-adult) molt. Sex-specific differences derive from two sources (1) the M-lineage (MaleMusFIG 3), which generates 41 sex muscles not present in hermaphrodites and (2) male-specific modification of certain juvenile muscles that are common to both sexes.
2.1 Male-specific (M-derived) Muscles
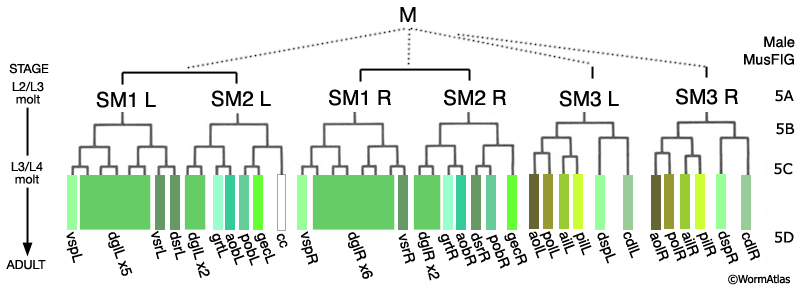
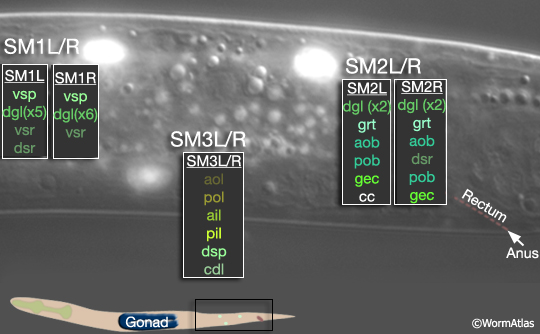
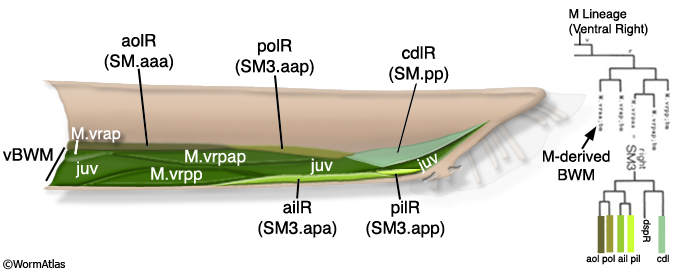
During late L2 the M lineage of the male executes a different lineage from that in the hermaphrodite and produces 3 bilateral pairs of Sex Myoblasts (SM1-3L/R) (MaleMusFIG 3 and 4). As in the hermaphrodite, SMs migrate from their birth position but move in the opposite direction, towards the tail instead of anteriorly (Sulston and Horvitz, 1977; Sulston et al., 1980; Branda and Stern, 2000). During L3 the SMs divide 2 or 3 times to generate 41 male-specific muscles (the sex muscles; MaleMusFIG 3, 5A-D). SM3L(left) and SM3R(right) have identical lineages, while the SM1left/right homologs as well as those of SM2 produce non-identical lineages (summarized in MaleMusFIG 4). Lineage execution is accompanied by a number of short-range migrations (see Sulston et al., 1980, Fig 27, for details).
MaleMusFIG 3: The M lineage. (L) Left; (R) right; (ail and pil) anterior and posterior inner longitudinal; (aob and pob) anterior and posterior oblique; (aol and pol) anterior and posterior longitudinal; (cc) coelomocyte; (cdl) caudal outer longitudinal; (dgl) diagonal; (gec) gubernacular erector; (grt) gubernacular retractor; (vsr and dsr) ventral and dorsal spicule retractors; (vsp and dsp) ventral and spicule protractors.
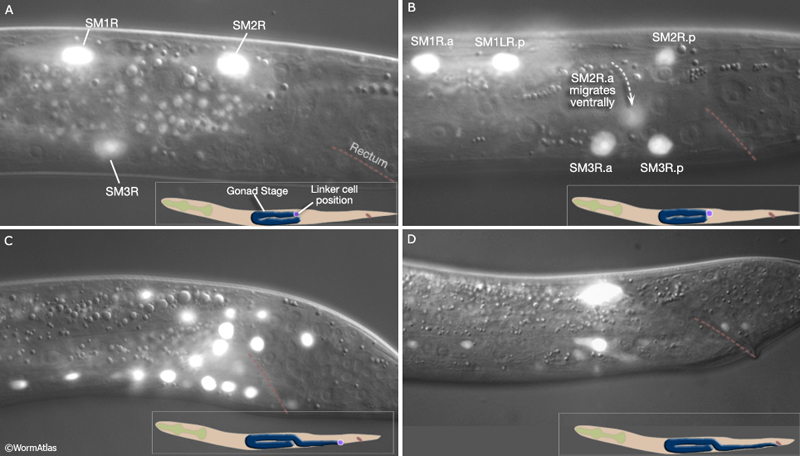
MaleMusFIG 4: Muscles produced by the SMs. DIC/ epifluorescent image from transgenic L3 animal expressing the hlh-8 ::GFP reporter gene (transcription factor). (Strain source: B.D. Harfe, M. Krause and A. Fire.)
|
MaleMusFIG 5: Division of the SMs to produce the sex muscles. Epifluorescent images from transgenic animals expressing the hlh-8 ::GFP reporter gene (transcription factor). (Strain source: B.D. Harfe, M. Krause and A. Fire.) A. L3 animal at approximately 27 hours, lateral view, right side. B. L3 animal at approximately 30 hours, lateral view, right side. C. L4 animal at approximately 35 hours, lateral view, right side. Terminal cells are born (SM.xx/xxx). Several migrate to reach final position (aob,pob, grt, gec). D. L4 animal during morphogenesis at approximately 42 hours, lateral view, left side. Terminal cells in final position. (Fewer cells express reporter at this stage as HLH-8::GFP signal diminishes in maturing terminal cells.)
2.2 Sexually Dimorphic Muscles
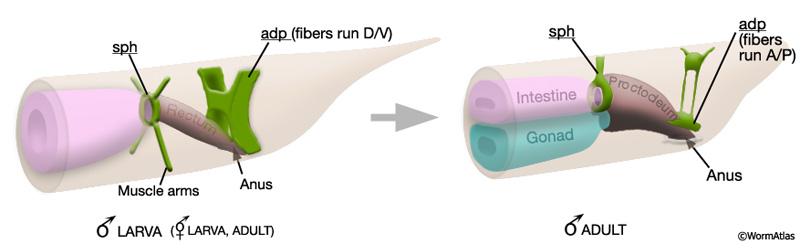
Some tail muscles that are present in larvae of both sexes become modified in the male late in development to function in mating behavior or copulation. These alterations can be subtle, such as the establishment of connections between non-sex-specific muscles and male-specific muscles or neurons. For example, dorsal body wall muscles, present in both sexes, can be considered sexually dimorphic because they establish gap junctions with spicule retractor muscles in the male (Male-specific Muscles; MaleMusFIG25 C). Other modifications are more striking, for example, the anal depressor (adp) and the sphincter (sph) undergo extensive morphological and functional changes in the male but not in the hermaphrodite (Sulston et al., 1980).
The male anal depressor (adp): In both male and hermaphrodite larvae the adp is an H-shaped cell connecting the dorsal body wall to the roof of the rectum (MaleMusFIG 6). Muscle myofilaments run dorsoventrally (D/V) and their contraction lifts the roof of the anus to allow defecation (MaleMusFIG 7). During male L4 lethargus, the adp substantially changes its shape and its contacts. The cell tilts forwards slightly but does not completely lose its contact with the dorsal roof (MaleMusFIG 6). Ventrally, the anterior edges of the cell grow forward so that they meet the dorsal spicule protractor muscles and form attachments with these cells (described in Male-specific Muscles). The myofilaments within the muscle reorient so that they now run anteroposteriorly (A/P) (MaleMusFIG 8), the same direction as the spicule protractor (Sulston and Horvitz, 1977; Sulston et al., 1980). Thus the adp becomes an extension of the dorsal spicule protractors.
MaleMusFIG 6: The male anal depressor. Diagram showing the H-shaped anal depressor cell in larval and adult animals.
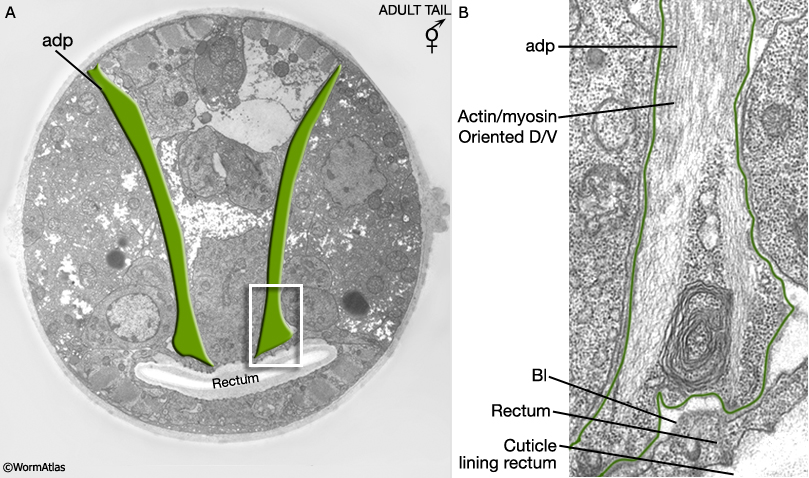
MaleMusFIG 7: Hermaphrodite anal depressor muscle. A. Electron micrograph featuring the anal depressor (adp), transverse view. (Image source: N2 [Hall] 550-52.) B. Detailed view of boxed region in A.(Bl) Basal lamina.
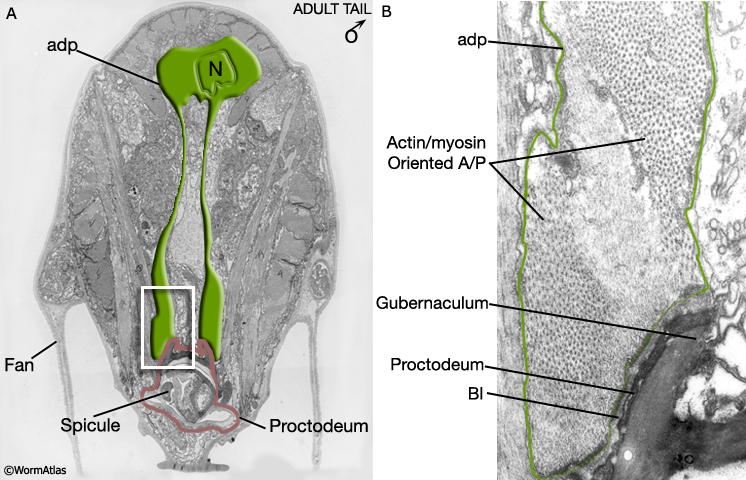
MaleMusFIG 8: Male anal depressor muscle. A. Electron micrograph featuring the anal depressor (adp), transverse view. (N) nucleus. (Image source: N2Y [MRC] 32.) B. Detailed view of boxed region in A. (Bl) Basal lamina.
The male sphincter (sph): Late in male development the sphincter is functionally and morphologically altered to compensate for a newly formed connection between the alimentary and reproductive tracts (MaleMusFIG 6; Sulston et al., 1980). The muscle becomes enlarged (hypertrophied) and sends an extension dorsally which attaches to the body wall roof. Intrinsic activity of the muscle and its response to GABA, which regulates defecation, are also altered (Reiner and Thomas, 1995; see Male-specific Muscles).
3 Diagonal Muscles
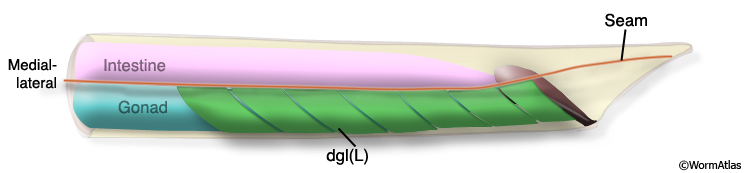
The adult male has 15 diagonal muscles (dgl's) organized in a herring-bone-like fashion along either side of the ventral midline; 8 muscles on the right side (dglR), 7 on the left (dglL) (MaleMusFIG 9 and 10A; Sulston et al., 1980).
MaleMusFIG 9: Diagonal muscles. Adult male, left lateral view.
|
|
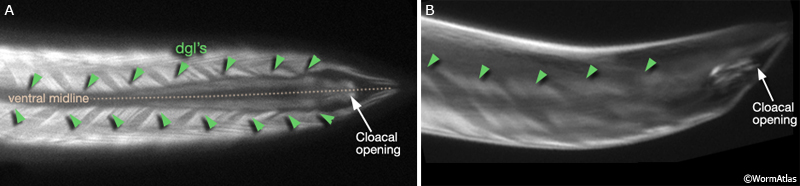
MaleMusFIG 10: Epifluorescent views of the diagonal muscles. Epifluorescent images from transgenic animals expressing the unc-27 reporter gene (Troponinl, sarcomere thin filaments). (Strain source: L. Jia and S.W. Emmons.) A. Ventral view. B. Left, lateral view.
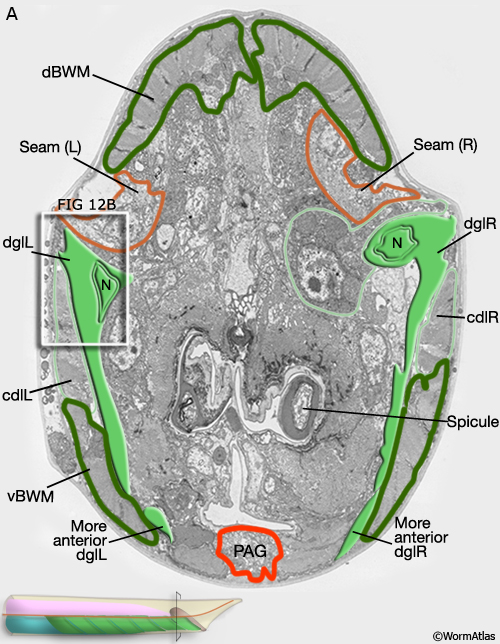
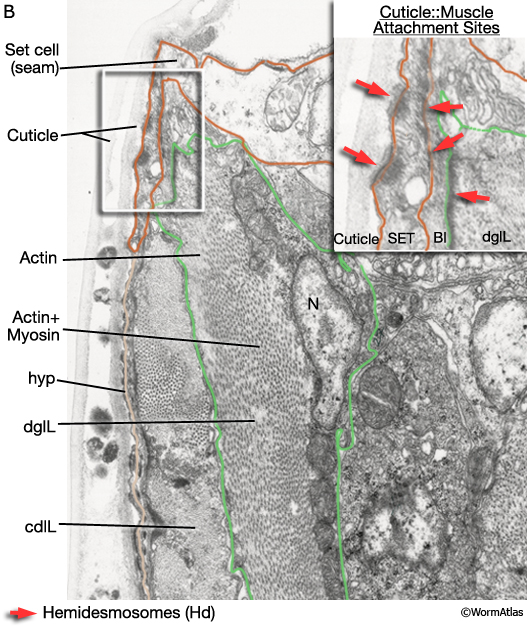
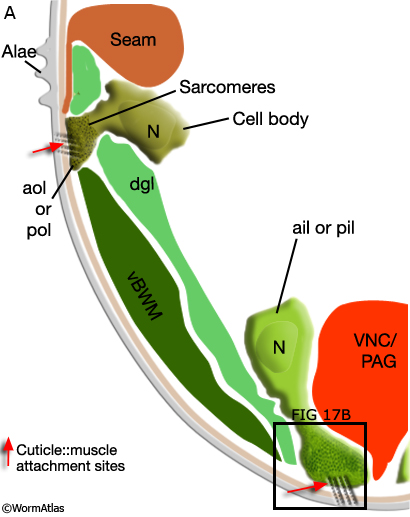
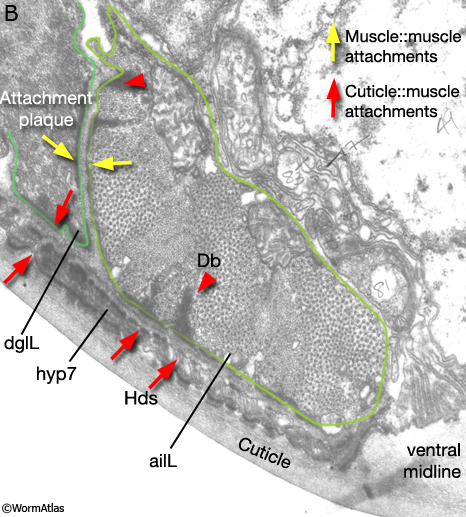
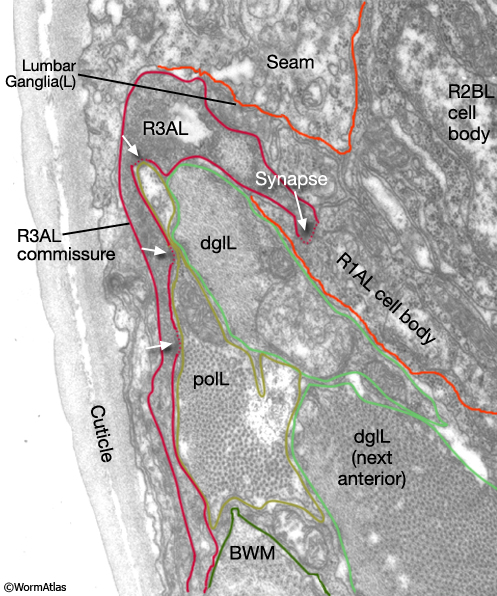
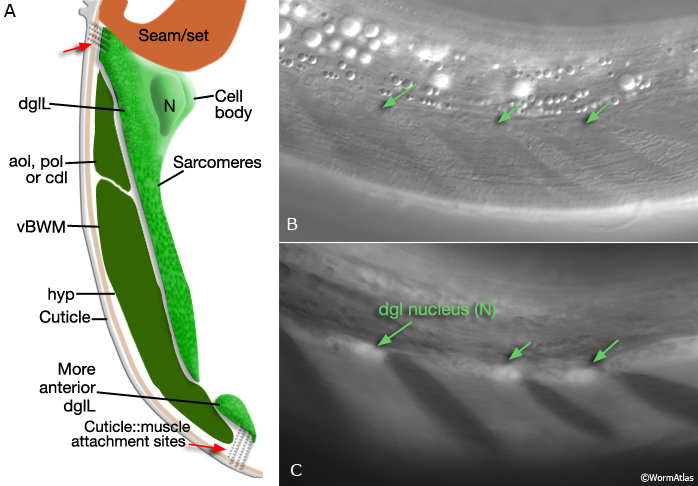
Viewed by polarized light or with muscle myofilament markers, the dgl's appear to contain 3 sarcomeres (MaleMusFIG 10B; Waterston, 1988). However, the myofilament banding patterning observed in electron micrographs suggests possibly fewer. Along their dorsal edges, the muscles attach to the body wall at the lateral seam and along their ventral edge near the ventral midline (MaleMusFIG 11 and 12B). The attachment points are localized at the ends of the muscle cells as half I bands ending in hemidesmosomes (Hd) and connect the myofilaments to basal lamina (Bl). The basal lamina is connected to Hds in the hyp and the hyp in turn to cuticle (red arrows, MaleMusFIG 11A, 12A and 12B; see Muscle System - Somatic Muscle for a detailed description of such structures). Diagonal muscles are also connected to other muscles, namely the inner longitudinal muscles (ailL/R, pilL/R). The muscles are joined via attachment plaques (muscle::muscle attachment complexes - yellow arrows in MaleMusFIG 17B, for attachment plaque description see Muscle System - Somatic Muscle).
MaleMusFIG 12A: Ultrastructure of the diagonal muscles. Low-power TEM showing the relationship of the diagonal muscle with the seam, transverse section. (PAG) Pre-anal ganglion. (Image source: N2Y [MRC] 579-20.)
MaleMusFIG 12B: Ultrastructure of the adult diagonal muscle. Higher magnification TEM of region highlighted by white box in 12A featuring diagonal muscle with actin and myosin filaments, transverse view. Inset shows cuticle to muscle attachment sites. (Red arrows) Hemidesmosomes; (Bl) basal lamina; (SET) set cell. (Image source: N2Y [MRC] 550-4.)
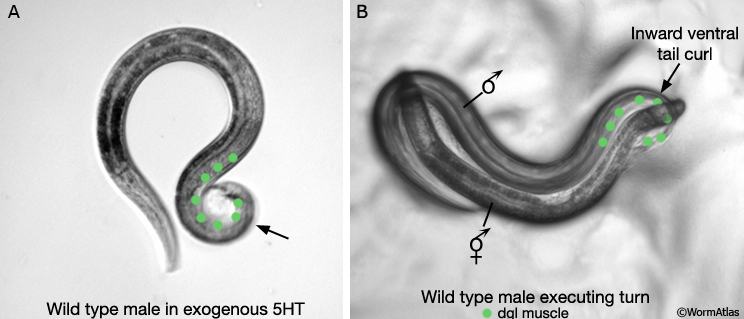
During mating when males reach the end of the hermaphrodite they execute a turn to flip to the other side (MaleMusFIG 13B; MaleMusMOVIE 1). This turn requires ventral curling of the tail facilitated by contraction of diagonal and other tail muscles. The diagonal muscles are innervated by the male-specific CP motor neurons of the ventral cord (White, 1988). These neurons are required for precise execution of the turn (Loer and Kenyon, 1993). Serotonin (5HT) produced by the CP neurons may be responsible for stimulating diagonal muscle contraction during turning as exposure of males to exogenous serotonin can induce ventral tail curling (MaleMusFIG 13A).
MaleMusFIG 13: Male turning behavior. A. Exposure of males to exogenous serotonin (5HT) induces ventral tail curling. B. Ventral curling of male tail when the male reaches the end of the hermaphrodite body during mating.
4 Longitudinal Muscles
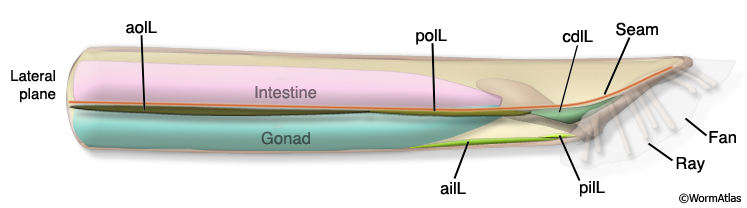
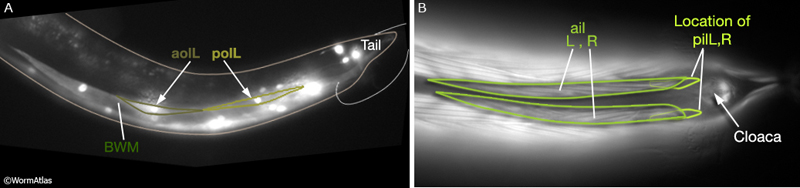
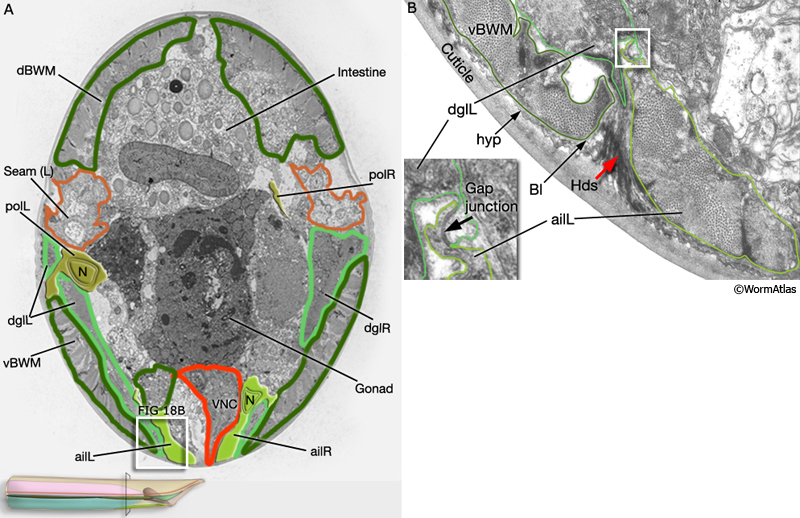
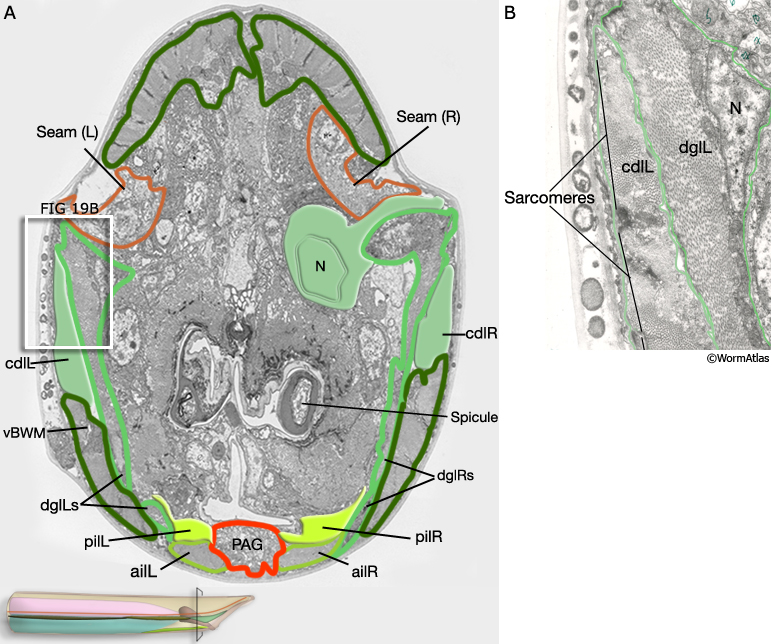
Three sets of longitudinal muscles are generated by the M lineage and contribute to the body wall musculature of the male (Sulston et al., 1980; MaleMusFIG 14 and 15): (1) the anterior (aolL/R) and posterior (polL/R) OUTER longitudinal muscles, positioned just below the seam (MaleMusFIG 16,18A and 19A); (2) the caudal OUTER longitudinal muscles (cdlL/R), which replace pol(L/R) in the tail (MaleMusFIG 19A); (3) the anterior (ailL/R) and posterior (pilL/R) INNER longitudinal muscles, positioned on either side of the ventral midline (MaleMusFIG 16 and 19A).
MaleMusFIG 14: Longitudinal muscles. Illustration of adult male longitudinal muscles, left lateral side.
MaleMusFIG 15: Relationship between longitudinal and body wall muscles. Diagram of physical and lineal relationship between longitudinal muscles and ventral body wall muscles (vBWM), right side. BWM are derived from MS. (Based on Sulston et al., 1980.)
In contrast to other sex muscles of the male which are non-striated, the longitudinal muscles, particularly ail(L/R), pil(L/R) and cdl(L/R), are striated, like their body wall muscle neighbors (BWM) (Sulston J.E., Albertson, D.G. and Thomson J. N., unpublished). They contain significantly more sarcomeres than non-striated muscles and attach to the body wall cuticle at multiple sites along their length (in contrast to non-striated muscles which attach only at their ends; MaleMusFIG 16, 17, 18B, 19B and 20).
MaleMusFIG 16: Sarcomeres of the longitudinal muscles. A. Epifluorescent image from transgenic animal expressing the mab-23 reporter gene (transcription factor), left lateral view. (Strain source: R. Lints and S.W. Emmons.) B. Epifluorescent image from transgenic animal expressing the unc-27 reporter gene (Troponinl), ventral view. (Strain source: L. Jia and S.W. Emmons.)
MaleMusFIG 17A: Longitudinal muscles. Diagram, transverse view. Basal laminae not shown.
MaleMusFIG 17B: Ultrastructure of longitudinal muscle. TEM, transverse view of boxed area in 17A. Image shows attachment sites of muscle to cuticle (red arrows) and muscle to muscle (yellow arrows). (Image source: N2Y [MRC] 1024.)
The ails and pils are joined via attachment plaques and by gap junctions to diagonal muscles (Sulston J.E., Albertson, D.G. and Thomson J. N., unpublished). The longitudinal muscles may therefore function with diagonal muscles in turning (MaleMusFIG 18B, 19B). Consistent with this possibility, the posterior outer longitudinal muscles receive inputs from the rays, which have been shown to play a role in turning (MaleMusFIG 20; Liu and Sternberg, 1995). These NMJs occur on the muscle sarcomere, and not on muscle arms, as the neuron enters the commissures to the preanal ganglion.
MaleMusFIG 18: Anterior and posterior longitudinal muscles. A. Low-power TEM showing the relationship of the longitudinal muscle with the diagonal and body wall muscles, transverse section. B. TEM, transverse view of boxed area in 18A. (Red arrow) Hemidesmosome. (VNC) Ventral nerve cord; (vBWM) ventral body wall muscle. (Image source: N2Y [MRC] 595-19.)
MaleMusFIG 19: Caudal longitudinal muscles. A. Low-power TEM showing the relationship of the caudal longitudinal muscle with the diagonal muscles, transverse section. B. TEM, higher magnification of boxed area in 19A. (N) Nucleus; (PAG) pre-anal ganglion; (vBWM) ventral body wall muscle. (Image source: N2Y [MRC] 579-20.)
MaleMusFIG 20: NMJs of longitudinal muscles. TEM showing synapses between longitudinal muscle and R3AL commissure from the rays, transverse view.(polL) Posterior outer longitudinal; (dglL) diagonal muscle; (BWM) body wall muscle. (Image source: N2Y [MRC] L849.)
5 References
Barr, M.M. and Sternberg, P.W. 1999. A polycystic kidney-disease gene homologue required for male mating behaviour in C. elegans. Nature 401: 339-40. Abstract
Branda, C.S. and Stern, M.J. 2000. Mechanisms controlling sex myoblast migration in Caenorhabditis elegans hermaphrodites. Dev. Biol. 226: 137-51. Article
Garcia, L.R., Mehta, P. and Sternberg, P.W. 2001. Regulation of distinct muscle behaviors controls the C. elegans male's copulatory spicules during mating. Cell 107: 777-788. Article
Liu, K.S. and Sternberg, P.W. 1995. Sensory regulation of male mating behavior in Caenorhabditis elegans. Neuron 14: 79-89 Article
Loer, C.M. and Kenyon, C.J. 1993. Serotonin-deficient mutants and male mating behavior in the nematode Caenorhabditis elegans. J. Neurosci. 13: 5407-17. Article
Reiner, D.J. and Thomas, J.H. 1995. Reversal of a muscle response to GABA during C. elegans male development. J. Neurosci. 15: 6094-6102. Article
Sulston, J.E. and Horvitz, H. R. 1977. Post-embryonic cell lineages of the nematode Caenorhabditis elegans. Dev. Biol. 56: 110-156. Article
Sulston, J.E., Albertson, D.G. and Thomson, J.N. 1980. The Caenorhabditis elegans male: Postembryonic development of nongonadal structures. Dev Biol. 78: 542-576. Article
Sulston, J.E., Schierenberg, E., White J.G. and Thomson, J.N. 1983. The embryonic cell lineage of the nematode Caenorhabditis elegans. Dev. Biol. 100: 64-119. Article
Waterston, R.H. 1988. Muscle. In The nematode C. elegans (ed. W.B. Wood). Chapter 10. pp 281-335. Cold Spring Harbor Laboratory Press, Cold Spring Harbor, New York.
White, J. 1988. The Anatomy. In The nematode C. elegans (ed. W.B. Wood). Chapter 4. pp 81-122. Cold Spring Harbor Laboratory Press, Cold Spring Harbor, New York.
|

This chapter should be cited as: Lints, R. and Hall, D.H. 2009. Male muscle system, overview. In WormAtlas. doi:10.3908/wormatlas.2.4
Edited for the web by Laura A. Herndon. Last revision: July 11, 2013. |
|