3 Rectal Gland
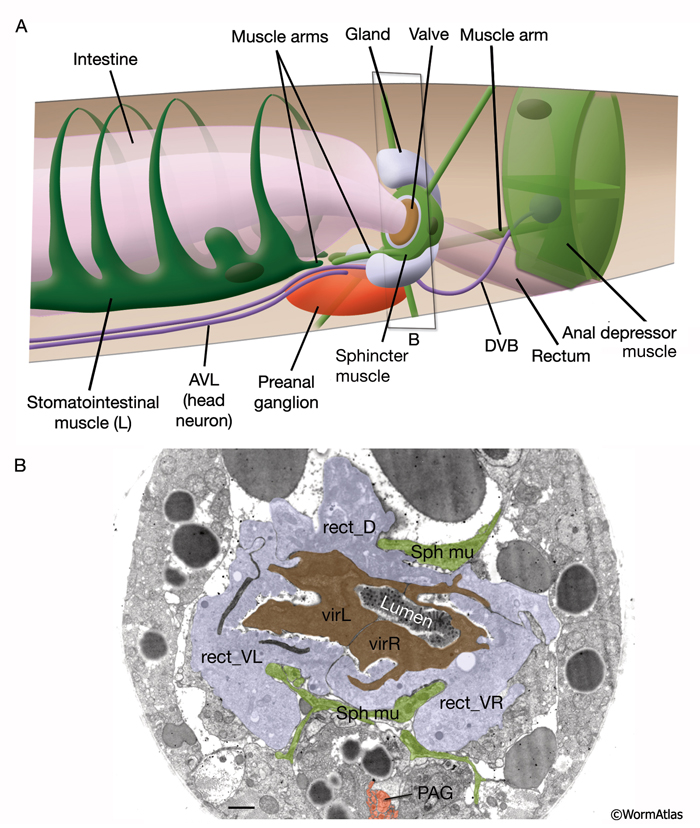
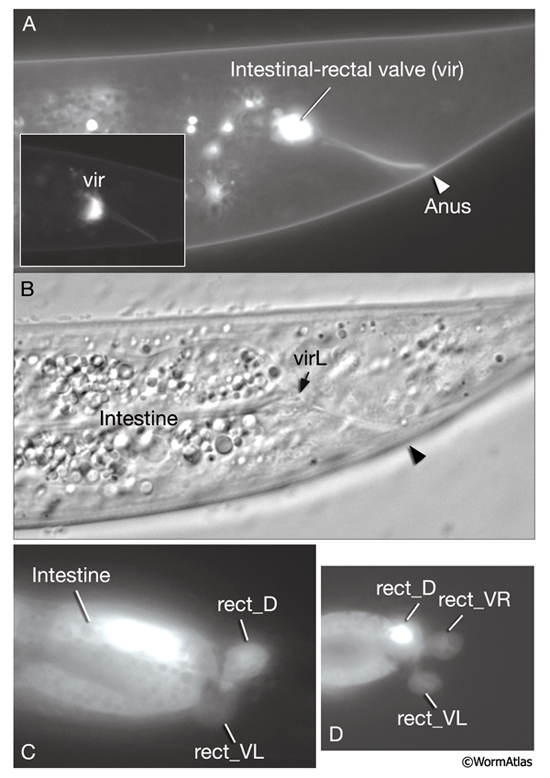
A ring of three large rectal gland cells - rect_D, rect_VL, rect_VR (in some older literature, these cells are also referred to as “rep”) - connect to the intestinal lumen just posterior to the rectal valve (RectFIG 2). It is possible that these cells secrete digestive enzymes into the caudal lumen of the intestine, which is slightly inflated compared to lumen in the midbody. The cells lie at the same level or just behind the rectal valve, and their apical specialization facing the lumen produces both microvilli (similar to intestinal cells) and cuticle (similar to transitional epithelia) in discrete patches (D.H. Hall, unpubl.).
4 Rectal Epithelium
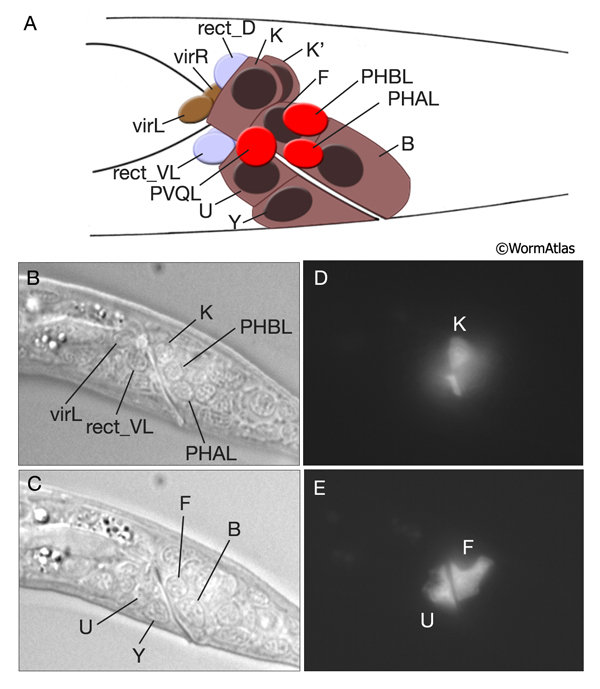
Six cells of the posterior hindgut - B, F, Y (formerly called post-embryonic C), U (formerly called post-embryonic E), K and K’ - are rectal epithelial cells (IntFIG 2 and RectFIG 3). Each produces some portion of the cuticular lining of the rectum, and they all contain secretory membrane stacks along the region facing the lumenal cuticle. Their cytoplasmic contents otherwise resemble that of typical hypodermal cells. These cells connect the intestinal-rectal valve to the main body hypodermal syncytium (hyp 7). In the hermaphrodite, the Y cell becomes the PDA neuron in a cell fate switch after early larval stages. Also, at late L1, the K cell divides to generate the Ka daughter as the new rectal epithelial cell, whereas the Kp daughter becomes the DVB neuron. B, F, U and Y act as post-embryonic blast cells in the male.
RectFIG 3A-E: Development of the rectum. A. Graphic rendition of rectal cells in early L1 larva. Their nuclei are shown as dark ovals. Nuclei of some of the neurons in this region, the gland cells and the valve cells are also shown. The rectal epithelial K and K' cells lie as a left and right pair, whereas the remaining rectal epithelial cells, B, F, U and Y are placed dorso-ventrally around the lumen. (Based on Sulston et al, 1983). B-E. Rectal epithelial cells in L1 larva. (B, C) DIC images, left lateral views. B is taken from a plane more lateral than C and shows the nucleus of K as well as the neuron nuclei. Four other rectal nuclei are seen at a medial plane in C. (D, E) Epifluorescent images from the same animal expressing the transgene lin-48::GFP in some rectal epithelial cells. (Strain Source: H. Chamberlin.) Magnification, 600x.
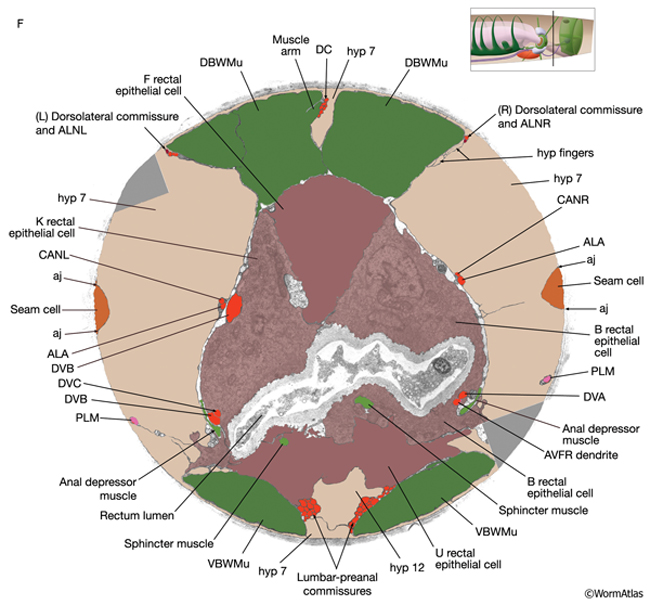
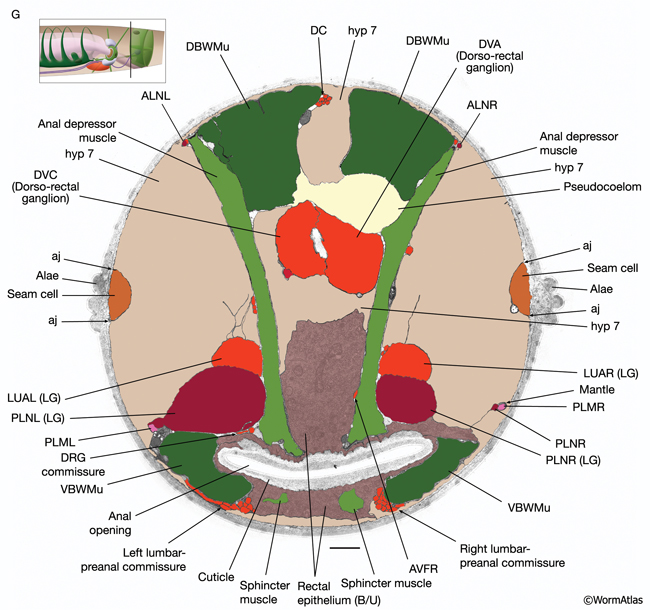
RectFIG 3F&G: Transverse pseudocolored TEM images of the adult rectum. The K cell has divided and the Y cell has become PDA (not shown). Hyp 12 becomes part of hyp7. (DC) Dorsal cord; (LG) lumbar ganglia; (aj) adherens junction; (DBWMu and VBWMu) dorsal and ventral body wall muscles, respectively. F shows the middle of the rectum, whereas G is closer to the anal opening. (Image source: [Hall] B140B [dark gray areas are gaps in the original TEM montage]. Bar, 1 μm.
The Pax transcription factor EGL-38 has been found to be important for the development of hindgut cell types. Downstream of EGL-38, a combination of transcription factors contribute to each cell’s fate (Chamberlin et al., 1997; Sewell et al., 2003) |
5 Muscle Cells of the Hindgut (Enteric Muscles)
The four specialized muscle cells of the hindgut (the two stomatointestinal muscles [also called the intestinal muscles], the anal sphincter muscle [also known as the anal dilator or rectal muscle], and the anal depressor muscle [also called the depressor ani muscle]), are reviewed in detail in the Muscle System - Nonstriated. These enteric muscles operate jointly in the defecation cycle. The sphincter and anal depressor muscles are anchored to the body wall and to the rectal epithelium (RectFIG 3). All four muscles send arms to the DVB neuron along dorsal surface of the preanal ganglion. The DVB neuron makes synapses onto the arms of stomatointestinal muscle and the anal depressor muscle. All three sets of muscles are coupled to each other via gap junctions (White et al., 1986) (see also Gap Junctions). Their coupled contractions control the enteric muscle contraction (EMC) step of defecation.
6 Motor Neurons of Defecation
In C. elegans, defecation is achieved through rhythmic activation of a stereotyped cycle of muscle contractions (Liu and Thomas, 1994). Laser ablation of the AVL and DVB neurons together eliminate enteric muscle contractions. AVL is an excitatory γ-aminobutyric acid (GABA)ergic interneuron/motorneuron that is located in the head and sends a process to the tail. It influences the enteric muscles indirectly via gap junctions with DVB (see also Gap Junctions). Killing AVL alone causes a strong anterior body muscle contraction (aBoc)-defective defecation phenotype. Because this defect is not seen in mutants that lack GABA, AVL may also use a non-GABAergic signal to activate other motor neurons that control anterior body wall muscle contraction. DVB is a GABAergic motor neuron located in the dorsorectal ganglion in tail (NeuroFIG 18). It is born post-embryonically at late L1 stage and is the daughter of the K rectal epithelial cell. DVB extends a prominent process anteriorly that passes through a commissure beneath the depressor muscle and extends forward along the top of the ventral hypodermal ridge. In this region, its large axon is filled with synaptic vesicles and makes periodic synapses to muscle arms from enteric muscles. The excitatory GABAergic signal from AVL and DVB is thought to be mediated by the nonselective cation-channel-type GABA receptor EXP-1 because exp-1 mutants lack enteric muscle contractions and are phenotypically constipated (Thomas, 1990; Beg and Jorgensen, 2003).
7 Defecation Motor Program
In C. elegans, defecation consistently occurs approximately every 50 seconds and has five cycle components: an intercycle period, pBoc (posterior body muscle contraction), pBoc relaxation, aBoc and EMC (enteric muscle contraction) which is also called the expulsion (exp) step (Avery and Thomas, 1997). In the hermaphrodite, each defecation starts with pBoc, which squeezes intestinal contents anteriorly. Approximately 1 second later, relaxation occurs and intestinal contents flow posteriorly. Next, aBoc is initiated by contraction of the body muscles near the head, and gut contents are concentrated near the anus. Finally, contraction of the enteric muscles expels the gut contents out of the animal and the intercycle period starts. The motor components of defecation behavior (pBoc, aBoc and EMC) constitute the defecation motor program (DMP). Defects in any of the motor components of DMP lead to constipation. During larval stages the hindgut structures of the male are virtually identical to those of the hermaphrodite. In the adult male although DMP is similar to the hermaphrodite, the anatomy and control of the hindgut changes drastically (see Male - Defecation Muscles, Male Muscles - Overview).
The steps of the DMP are coordinated in precise temporal and spatial sequences. The ultradian defecation rhythm can be reset by light touch stimulus to the body and is thought to be controlled by an intestinal pacemaker that keeps time and activates the posterior body contraction at the start of each cycle (Dal Santo et al., 1999; Siklos et al., 2000). An essential element of this pattern generator and time keeper is the periodic, autonomous calcium release mediated by the inositol trisphosphate (IP3) receptor ITR-1 in the posterior intestine (Dal Santo et al., 1999). Intestinal calcium levels oscillate with the same periodicity as the defecation cycle and reach their peak levels just prior to the first muscle contraction (pBoc) step. Transient increases in calcium ions then propagate from the posterior to the anterior intestine (Espelt et al., 2005; Teramoto and Iwasaki, 2006). Blocking propagation of this calcium wave stops the later phases of the defecation motor program. In addition, mutations in itr-1 slow down or eliminate the cycle, further supporting the idea that IP3 receptor activity and calcium release rather than neuronal control sets the defecation cycle frequency. Nevertheless, the GABAergic motor neurons, AVL and DVB are required for the execution of the anterior body contraction and the enteric muscle contractions for expulsion (McIntire et al., 1993; Avery and Thomas, 1997). An intercellular signal originating from the calcium oscillations in the posterior intestine may activate these two neurons for later muscle contractions.
8 List of Cells of the Rectum and Anus
1. Intestinal-Rectal Valve Cells
virL
virR
2. Rectal Gland Cells
rectD
rectVL
rectVR
3. Rectal Epithelial Cells
B
F
Y
U
K'
K
K.a (Rect_hyp)
4. Enteric muscles
i. Stomatointestinal muscle
mu intL
mu intR
ii. Anal Sphincter muscle
mu sph
iii. Anal depressor muscle
mu anal
5. Enteric neurons
AVL
DVB
9 References
Avery D.G. and Thomas, J.H. 1997. Feeding and defecation. In C. elegans Volume II. Ed.s Riddle D.L., Blumenthal, T., Meyer B.J. and Priess J.R . Pp 679-716. Cold Spring Harbor Laboratory Press. Article
Beg, A.A. and Jorgensen, E.M. 2003. EXP-1 is an excitatory GABA-gated cation channel. Nature Neurosci. 6: 1145-1152. Abstract
Bowerman, B., Tax, F.E., Thomas, J.H. and Priess, J.R. 1992. Cell interactions involved in development of the bilaterally symmetrical intestinal valve cells during embryogenesis in Caenorhabditis elegans. Development 116: 1113-1122. Article
Bucher, E.A. and Seydoux, G.C. 1994. Gastrulation in the nematode Caenorhabditis elegans. Sem. Dev. Biol. 5: 121-130. Abstract
Chamberlin, H.M., Palmer, R.E., Newman, A.P., Sternberg, P.W., Baillie, D.L. and Thomas, J.H. 1997. The PAX gene egl-38 mediates developmental patterning in Caenorhabditis elegans. Development. 124: 3919-3928. Article
Dal Santo, P., Logan, M.A., Chisholm, A.D.,and Jorgensen, E.M. 1999. The inositol triphosphate receptor regulates a 50-second behavioral rhythm in C. elegans. Cell 98: 757-767. Article
Espelt, M.V., Estevez, A.Y., Yin, X. and Strange, K. 2005. Oscillatory Ca2+ signaling in the isolated Caenorhabditis elegans intestine: role of the inositol-1,4,5-trisphosphate receptor and phospholipases C beta and gamma. J. Gen. Physiol. 126: 379–392. Article
Liu, D.W.C. and Thomas, J.H. 1994. Regulation of a periodic motor program in C. elegans. J. Neurosci. 14: 1953-1962. Article
McIntire, S.L., Jorgensen, E., Kaplan, J. and Horvitz, H.R. 1993. The GABAergic nervous system of Caenorhabditis elegans. Nature. 364: 337-341. Abstract
Sewell, S.T., Zhang, G., Uttam, A. and Chamberlin, H.M. 2003. Developmental patterning in the Caenorhabditis elegans hindgut. Dev. Biol. 262: 88-93. Article
Siklos, S., Jasper, J.A., Wicks, S.R. and Rankin, C.H. 2000. Interactions between an endogenous oscillator and response to tap in C. elegans. Psychobiology 28: 571-580. Abstract
Sulston, J.E., Schierenberg, E., White, J.G. and Thomson, J.N. 1983. The embryonic cell lineage of the nematode Caenorhabditis elegans. Dev. Biol. 100: 64–119. Article
Teramoto, T. and Iwasaki, K. 2006. Intestinal calcium waves coordinate a behavioral motor program in C.elegans. Cell Calcium 40: 319–327. Abstract
Thomas, J.H. 1990. Genetic analysis of defecation in Caenorhabditis elegans. Genetics 124: 855-872. Article
White J.G., Southgate, E., Thomson, J.N. and Brenner, S. 1986. The structure of the nervous system of the nematode C. elegans. Philos. Trans. R. Soc. Lond. Series B. Biol. Sci. 314: 1-340. Article
|

 Click pictures for new window with figure and legend, click again for high resolution image
Click pictures for new window with figure and legend, click again for high resolution image