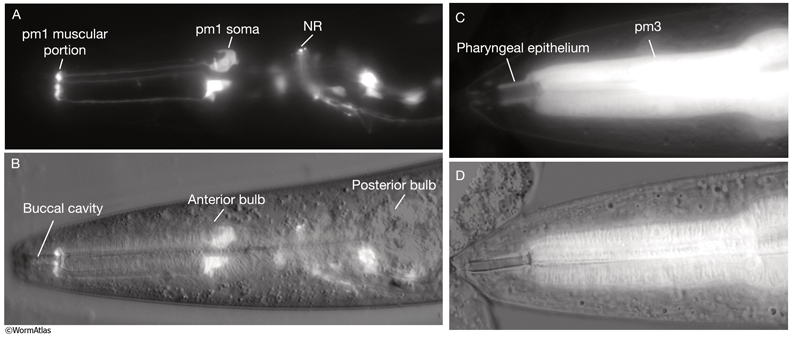
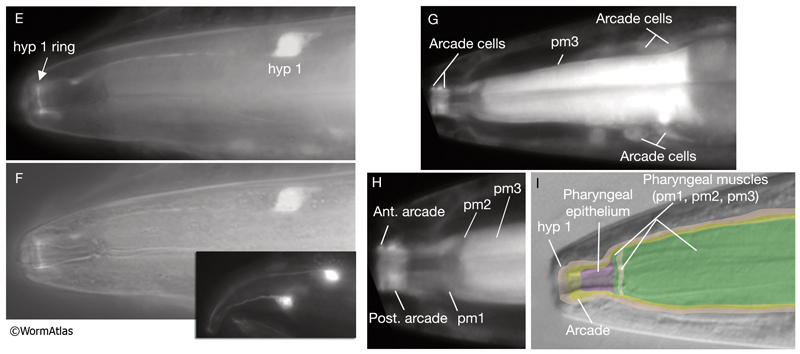
4 Muscle Cells of the Pharynx
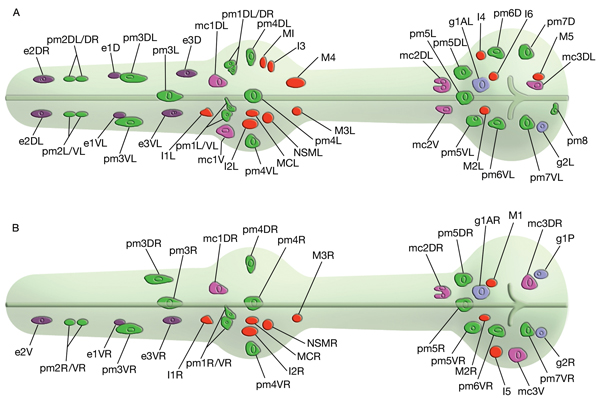
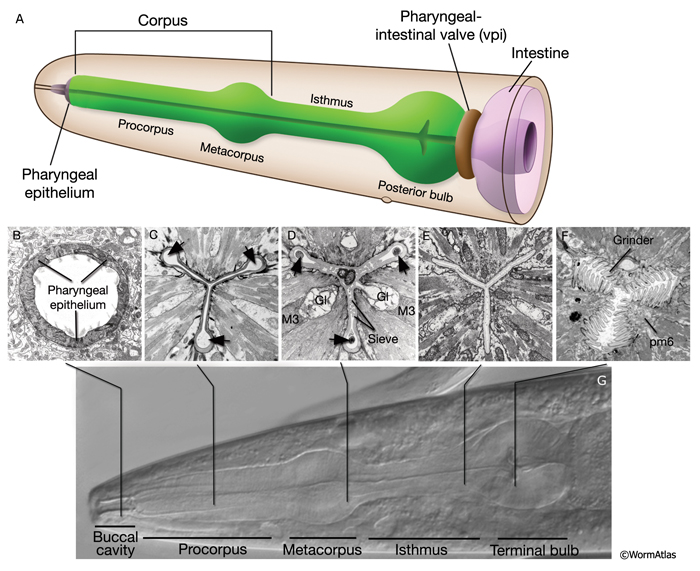
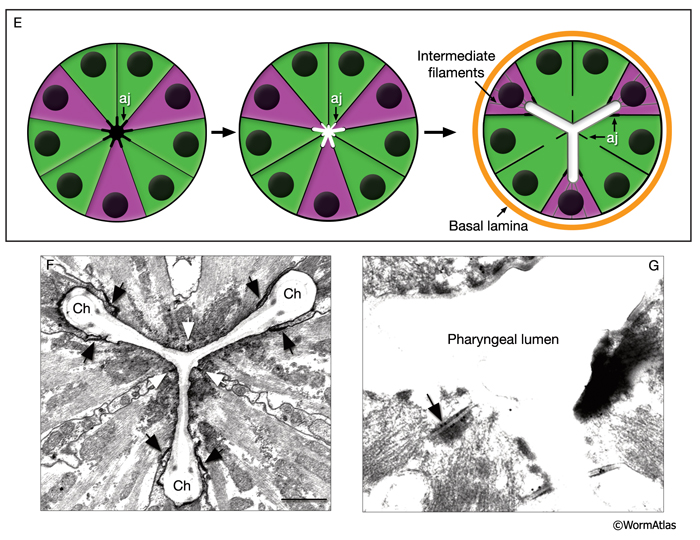
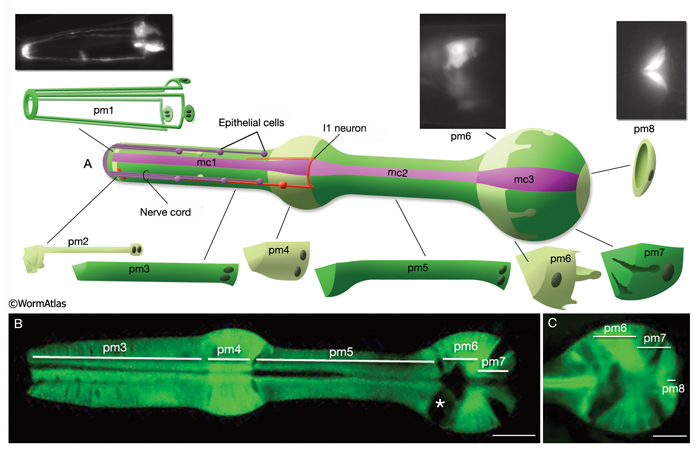
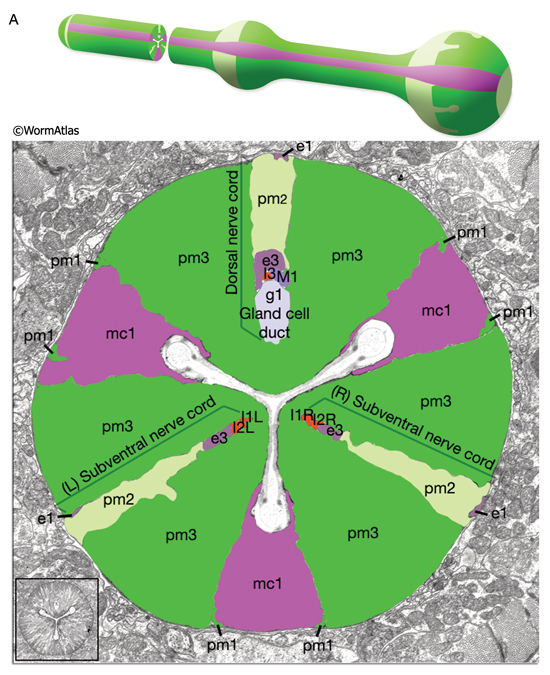
The pharyngeal muscles are grouped into eight separate segments (pm1-pm8), which are arranged as eight consecutive rings encircling the pharynx (PhaFIG 6;PharynxAtlas) (Albertson and Thomson 1976; Avery and Thomas, 1997; Franks et al., 2006). Unlike the body wall muscles, no hypodermal layer separates the muscle anchorage from the cuticle lining the lumen of the pharynx. Indeed, most of the pharyngeal muscles appear to participate directly in secreting cuticle (e.g., the two anteriormost pharyngeal muscle cells, pm1 and pm2, secrete the cuticle that lines the metostom and telostom portions of the buccal cavity) and thus qualify as having myoepithelial properties (Albertson and Thomson 1976, D.H. Hall, unpublished). Most of the pharyngeal muscle segments are made up of three syncytial cells positioned in a three-fold symmetrical manner in any cross section. Each of these cells contains two nuclei as a result of fusion of two cells around the time of hatching (See Table 1 in Albertson and Thomson 1976; see also PhaTABLE 1). The exceptions are pm1 and pm6-pm8; pm1 is a syncytial cell that contains six nuclei, pm6 and pm7 are made of three nonsyncytial cells each, and pm8 is a single, nonsyncytial cell. In pm2-pm5 of adult pharynx, there are still small remnant adherens junctions at the sites of former cell fusions between pairs of muscle cells, marking the exact apical border where the fusion has occurred (PhaFIG 3) (Hedgecock and Thomson, 1982). These remnant junctions express the AJM-1 protein and can be stained by immunocytochemistry (Koppen et al., 2001; D.H. Hall, unpublished). The three muscle cells of each segment are separated from each other by three marginal cells, whereas the muscle cells of the neighboring rings are linked by gap junctions and also connected via interlocking short fingers on the anterior and posterior margins. Each syncytial muscle cell contains a deep groove on the basal side where a longitudinal pharyngeal "nerve cord" is situated (PhaFIG 6 and PhaFIG 7A). These nerve cords enclose the cell bodies and processes of neurons, gland cells, and anterior epithelial cells. Many synapses, including neuromuscular junctions onto the pharyngeal muscles, occur along these cords.

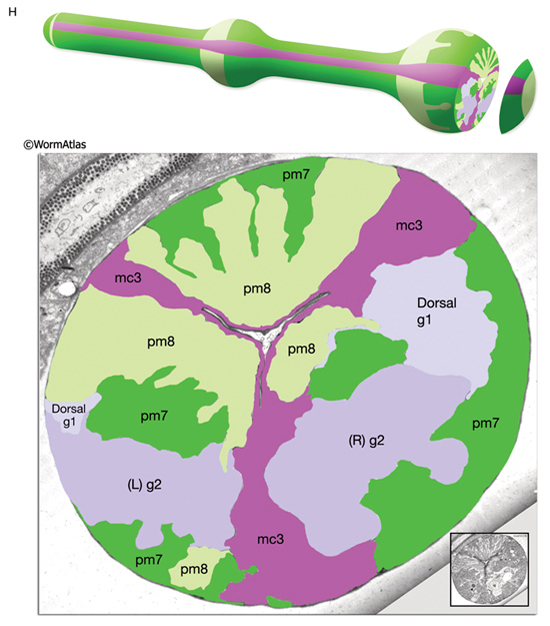
PhaFIG 6: Muscle cells of the pharynx. A. There are eight muscle segments (pm1-pm8) in the pharynx. All but pm6, pm7 and pm8 form syncytia. The most anterior segment, pm1, is a single cell with six nuclei. Its processes extend anteriorly adjacent to the outer edges of the marginal cells and terminate in a ring structure. Each pm2 soma has two nuclei and projects into a nerve ring to extend anteriorly. pm3 cells are located in procorpus, pm4 cells in the anterior bulb, and pm5 cells in isthmus. The terminal bulb contains pm6-pm8. The last muscle segment contains a single cell, pm8, that has a single nucleus located on the left side. (Top insets) Epifluorescent images of pm1 (left), pm6 (middle), and pm8 (right) in animals expressing GFP-tagged transgenes. (Strain source: Z-W. Wang and B. Chen.) Each magnification, 400x.
B. Pharyngeal muscle cells contain radially oriented filaments. Here, myofilaments in pm3-pm7 can be seen in an epifluorescent image taken from a transgenic animal expressing the reporter gene, C32F10.8::GFP. (Asterisk) Position of g1 gland cells. (Image source: R. Newbury.) C.
Muscles of the terminal bulb. Epifluorescent image from a transgenic animal expressing the reporter gene, K02A4.1::GFP. (Image source: R. Newbury.)
Specialized zones mark the apical regions of several of the pharyngeal muscles where they secrete cuticle. Here, the cytoplasm contains electron dense tubules, and sometimes, dense core vesicles just under the plasma membrane. The cytoplasm of pm6 is particularly specialized in the region underlying the grinder, which is a very elaborate cuticular structure secreted by the pm6 muscle cells (PhaFIG 2). Prominent networks of sarcoplasmic reticulum along the borders of each muscle sarcomere presumably sequester calcium needed for muscle contractility. Pharyngeal muscle cells also contain many mitochondria.
In most pharyngeal muscle segments, contractile filaments are oriented radially, and when the muscles contract, the pharyngeal lumen opens. However, in the terminal bulb, the pm7 muscle filaments are oriented obliquely with regard to the anterior-posterior axis and pull on the grinder region when the muscle contracts. pm1-pm4 function in sucking up and trapping bacteria. pm5 regulates flow of food from corpus to the terminal bulb, and pm6-pm8 operate the grinder.
5 Marginal Cells
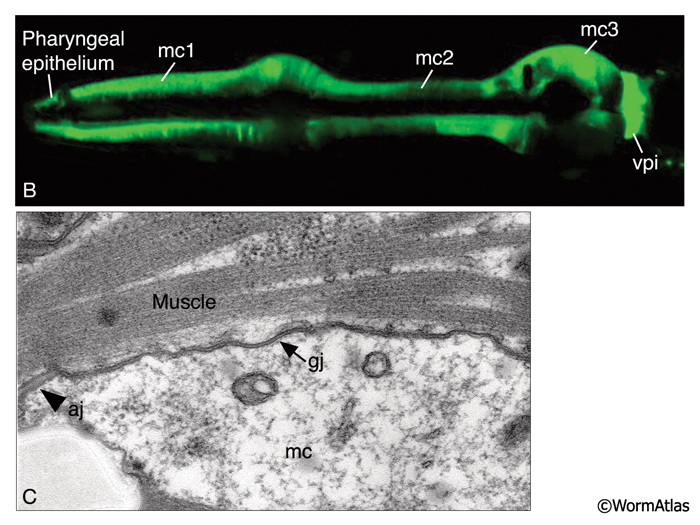
The pharynx is a mosaic of several non-equivalent cell types, each with threefold symmetry that assemble into a nonstratified, one-cell-deep epithelium along the lumen (PhaFIG 7; PharynxAtlas). As described above, muscle cells are a major part of this epithelium. Cells of another type, the marginal cells (mc), are placed at the three corners of the pharyngeal lumen and separate the muscle cells from one another. There are three mc segments along the pharynx and a total of seven marginal cells; three mc1 cells comprise the anterior segment, three mc2 cells comprise the second segment, and a syncytial mc3 cell with three nuclei comprises the terminal bulb segment (PhaFIG 1 and PhaFIG 6). From segment to segment, the marginal cells lie in rows at the corners of the lumen. They vary markedly in size between segments, but the three cells within one segment are essentially equivalent.
Marginal cells supply reinforcing strength to this muscular organ. Their large size and block-like shape is fortified by the placement of large radial bundles of intermediate filaments running from apical to basal borders of each cell. These filaments are anchored to the plasma membrane by large hemidesmosomes. Within each segment, the marginal cells are linked to neighboring muscle cells on their lateral borders by large gap junctions as well as apical adherens junctions separating the membrane to apical and basal surfaces (PhaFIG 7B&C) (Avery and Thomas, 1997). Large interlocking finger-like extensions also connect marginal cells to muscles within each segment and may also add to the structural integrity of the whole organ. Because of this arrangement, marginal cells within a segment communicate with muscle cells of the same segment and not with other marginal cells. In contrast, between segments, these cells form interlocking fingers and gap junctions to neighboring marginal cells.
The marginal cells contain many mitochondria, which suggests that these cells may perform some active role beyond merely providing continuity and strength to the epithelium. Because marginal cells are coupled to pharyngeal muscles via gap junctions, they may have some motor function, i.e., they may be myoepithelial in nature. Alternatively, they may act as relay stations to synchronously transmit signals from motor neurons to surrounding pharyngeal muscles so that all pharyngeal muscles within a segment can contract and relax at the same time. It is noteworthy that when the pharyngeal muscles contract, the muscle cells become thinner to open the lumen. Because the marginal cells are already relatively thin, this suggests that at full contraction, the pharyngeal lumen may open practically as wide as the inside corners of the marginal cells and form an open triangular lumen, whereas when the muscles relax, the lumen is practically closed except for the three channels at the apices of the anterior lumen.
6 Gland Cells of the Pharynx
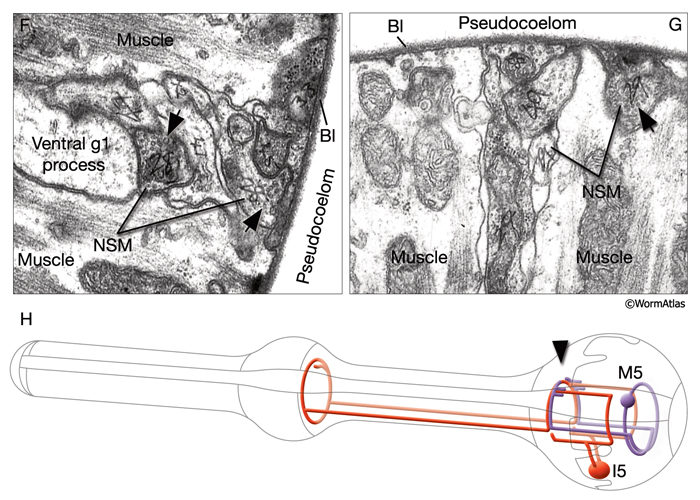
Two classes of gland cells, g1 (three cells) and g2 (two cells), are found in the second bulb of the pharynx (PhaFIG 8; PhaMOVIE 3; PharynxAtlas). Although previously dorsal g1 and right ventral g1 cells were reported to fuse, recent results with fluorescent marker expression studies suggest that these cells remain separate (Smit et al, 2008; J. Gaudet, pers. comm). The g1 cells extend three cuticle-lined ducts anteriorly within the narrow pharyngeal nerve cords. Two of these ducts pass through the isthmus before emptying into the pharyngeal lumen near the first bulb. The dorsal g1 duct travels much farther and empties near the anterior limit of the pharynx. The g2 cells also extend ducts, which are much shorter and empty into the lumen of the second bulb. The g1 cells contain a lamellar cytoplasm and few vesicles, whereas the g2 cells have a rather clear cytoplasm and more vesicles. These contents may vary from animal to animal, and vesicle sizes are quite large and variable. Gland cells receive motor innervation from M4 and M5 motor neurons, which suggests that they may be stimulated to secrete digestive enzymes synchronously with pharyngeal pumping activity. Periodic episodes of secretion (vesicle motion) have been seen in g1 ducts by light microscopy and are apparently associated with molting (Hall and Hedgecock, 1991; unpublished observations of J. Sulston as reported by Albertson and Thomson, 1976). This suggests that gland secretion may participate in the digestion of the pharyngeal cuticle during molting.
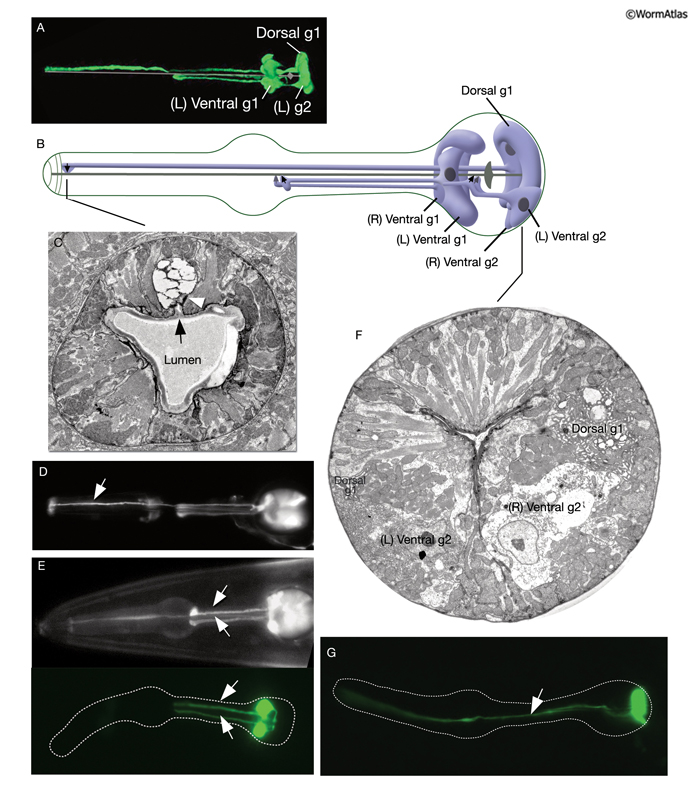
PhaMOVIE 3: 3-D reconstruction of pharyngeal gland cells. 3-D movie was created from confocal images of a strain expressing the GFP marker linked to the promoter for B0280.7 using Zeiss LSM 5 Pascal software v. 3.2. (Image source: R. Newbury and D. Moerman.) Click on image to play movie.
7 Pharyngeal Neurons
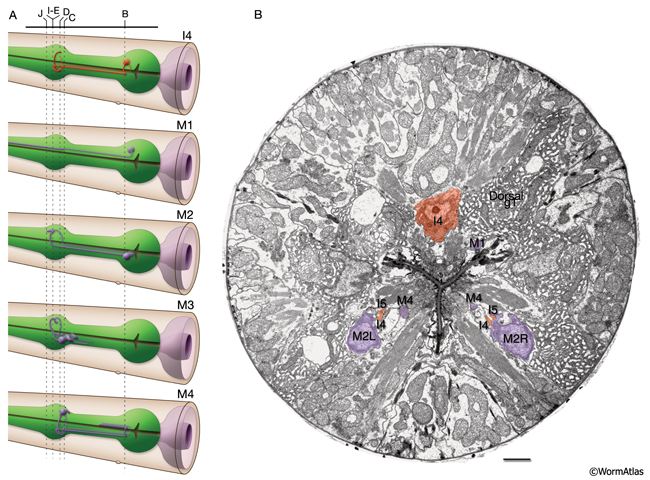
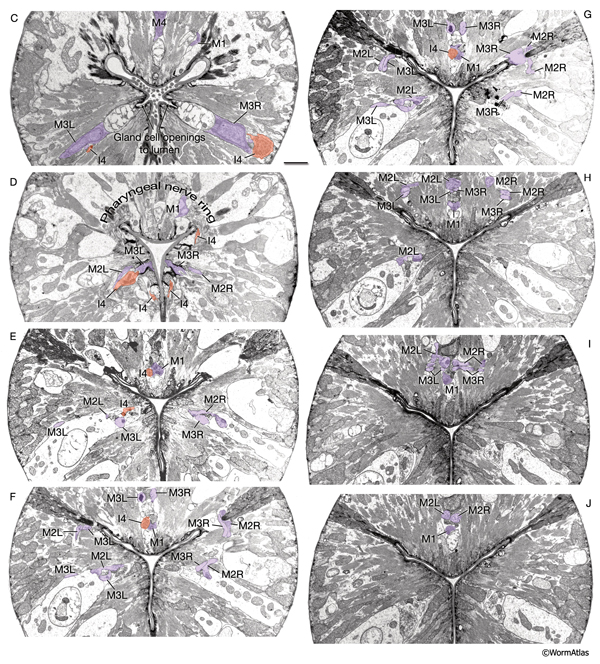
The pharynx has 20 intrinsic neurons of 14 types, all of which have cell bodies located in the anterior or posterior bulb (see also PharynxAtlas). Six types are bilaterally paired and eight are single neurons. These neurons extend processes anteriorly and/or posteriorly along the three longitudinal pharyngeal nerve cords (the dorsal and the right and left subventral) and form a small plexus (the pharyngeal nerve ring) within the anterior bulb where they decussate to the other side. A half-ring (the terminal bulb commissure) is made by neuronal processes more posteriorly within the anterior portion of the terminal bulb (PhaFIG 9 and PhaFIG 10). A few neurons (M1, M2 and M3) send out processes along unique routes. The M1 process runs anteriorly between the muscle and the right marginal cell until it reaches the pharyngeal nerve ring where it relocates to the dorsal nerve cord and continues traveling anteriorly in this location. Unlike other neuron processes, M2 and M3 processes do not make their dorsal turn within the pharyngeal nerve ring, but more anteriorly within the pm4 where they pierce through the pm4 as they travel towards the dorsal side to enter the dorsal nerve cord (PhaFIG 10E-J). M2 neurons make synapses to pm4 along the way.
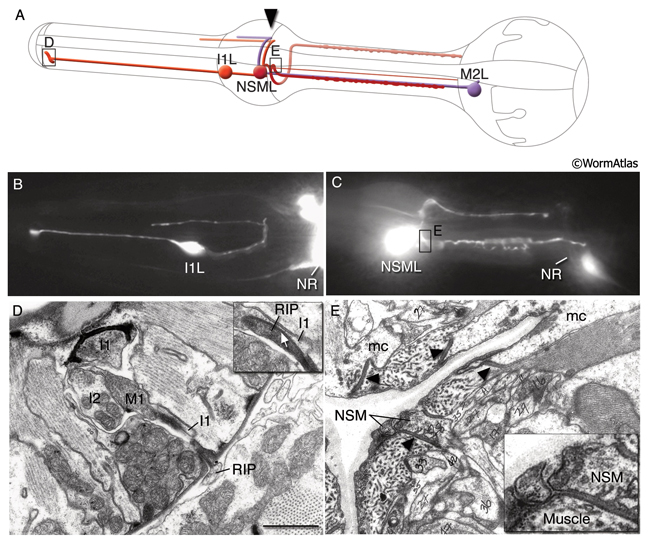
PhaFIG 10A&B: The processes of three of the pharyngeal neurons M1, M2 and M3. These neurons have unusual trajectories. All panels except A are transverse TEMs. Bars in B-J, 1 μm. (Image source: N2W [MRC] A382-A409.
A. Schematic illustration of section levels in panels B-J. B. Cell bodies of I4 and M2 neurons in the terminal bulb.
PhaFIG 10C-J: The processes of three of the pharyngeal neurons M1, M2 and M3. C. M3 cell bodies are near the ventral g1 duct openings into the lumen.
D. Pharyngeal nerve ring is composed of the circumferential neuron processes. E-J. Consecutive sections through the first bulb show the trajectories of M2 and M3 processes piercing pm4 to reach the dorsal cord.
Most of these processes interact synaptically with others within the pharyngeal nerve ring, formed between the narrow region where pm4 and pm5 appose each other (PhaFIG 9). These synapses are of the en passant type, similar to those seen in the somatic neurons. The pharyngeal motor neurons also form neuromuscular junctions (NMJs) onto pharyngeal muscles. Unlike NMJs between somatic neurons and body wall muscles, no basal lamina has been found separating the neurons from muscles in the pharynx. There are no direct contacts between pharyngeal neurons and the larger somatic nervous system except the gap junctions made between RIPL/R neurons and the pharyngeal I1s, and gap junctions between RIPL/R and the pharyngeal motor neuron, M1 (Albertson and Thomson, 1976; Avery and Thomas, 1997). Ablation of RIP cells results in only a minor effect on pharyngeal function such that the brief inhibition of pumping in response to light touch to body disappears (Avery and Thomas, 1997). It is possible that one or more pharyngeal neurons, such as NSM cells, could secrete hormonal factors into the pseudocoelom to influence the rest of the animal. Otherwise, pharyngeal and somatic nervous systems function more or less independently of each other. A dissected and isolated pharynx continues its normal pumping behavior.
In fact, generation of electrical potential changes required for pharyngeal pumping is probably intrinsic to the pharyngeal muscle cells themselves since pumping behavior continues even when all the pharyngeal neurons are ablated (Avery and Horvitz, 1989). Although pumping can still occur after complete ablation of the pharyngeal nervous system, four of the pharyngeal neurons, MC and M3 motor neuron pairs, are important for the regulation of the pump motion.
Pharyngeal neurons fall into three categories: motorneurons, interneurons and other neurons. However, this distinction is somewhat arbitrary because most of these neurons have structures that suggest mixed functions.
7.1 Pharyngeal Motor Neurons
Among motor neurons, M2 and M3 are paired neurons, whereas M1, M4 and M5 are single (PhaFIG 10; PharynxAtlas). M2 and M3 neurons are subventrally located and each innervates one side of the subventral and dorsal muscle sectors. M3s are also suggested to have proprioceptive sensory function. M4 and M5 neurons send out two branches, each of which first innervates the subventral sectors and then turns and innervates the dorsal sectors.
7.2 Interneurons
I1, I2, I3 and I6 are unbranched bipolar cells. I1 cells make electrical synapses with the somatic neurons RIPL/R and synapse onto MC neurons. I4 has two processes that grow anteriorly. I5 is a fairly complex cell, the processes of which make a circle within the pharyngeal nerve ring. All of these neurons except for I4 have free subcuticular endings that may have proprioceptive function. See PharynxAtlas for more details.
7.3 Other Neurons
MC neurons are a pair of bipolar cells situated within the anterior bulb that innervate pm4 (L. Avery pers. comm.). The motor-interneuron, MI, is a single unipolar cell situated in the dorsal side of the anterior bulb and synapses onto both muscle and other neurons. The neurosecretory-motor neurons NSML and NSMR send out branches that form varicosities and contain mixed-sized vesicles (PhaFIG 9). The two main processes of the NSMs run in close apposition to the pseudocoelom over most of their length and form some release sites towards this space. NSMs are serotonergic. These cells may have both neurosecretory and motor functions and may communicate the presence of food to the rest of the animal's body. Exogenous application of serotonin stimulates pumping, decreases locomotion and stimulates egg-laying. The same responses are seen in the presence of bacteria in the environment. NSMs were thought to be potential candidates for mediating the effects of endogenous serotonin. However, ablation of NSMss has only subtle effects on pumping, suggesting that they may be redundant for this function (Avery et al., 1993, Avery and Thomas, 1997). The slowing of locomotion in the presence of bacteria becomes more enhanced in animals that were previously food-deprived, compared to well-fed animals. This phenomenon is described as "the enhanced slowing response." When NSMs are ablated there is a small but significant decrease in this enhanced slowing of locomotion, which suggests NSMs contribute to this behavior (Sawin et al., 2000).
7.4 List of Pharyngeal Neurons
1. Motor neurons
M1
M2L/R
M3L/R
M4
M5
2. Interneurons
I1L/R
I2L/R
I3
I4
I5
I6
2. Other neurons
MI
NSML/R
MCL/R
See also PharynxAtlas
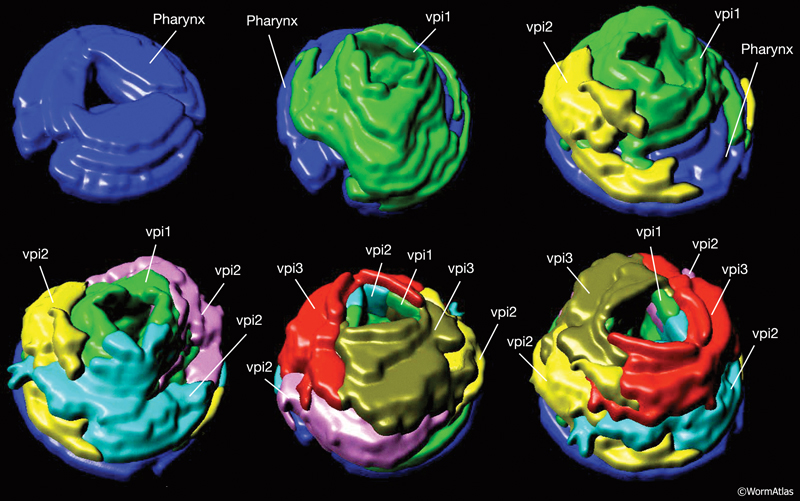
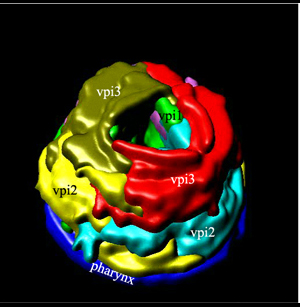
8 Pharyngeal-Intestinal Valve (VPI)
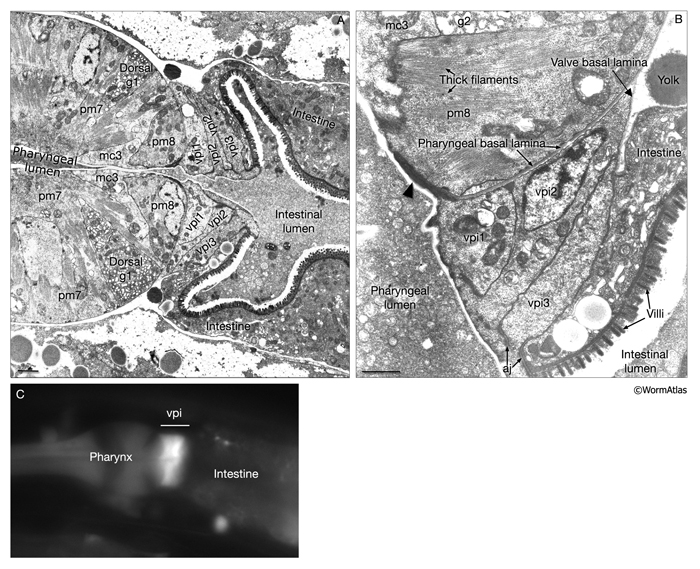
A group of six equivalent cells forms a tightly constructed “valve” that links the posterior bulb of the pharynx to the anterior four cells of the intestine (PhaFIG 11, PhaFIG 12 and PhaMOVIE 4.) These six cells comprise a small epithelial channel with a cuticular lining in continuity with the pharyngeal cuticle. The channel links the lumen of the pharynx to the large lumen of the anterior intestine. Three sets of cells form consecutive rings containing one, three and two cells from anterior to posterior (PhaFIG 12). The inside cuticle is reinforced by a series of closely spaced circumferential ridges, rather like the cuticle of the anterior buccal cavity. The valve cells are not syncytial, but are firmly linked to their neighbors and to the pharynx and/or intestine by robust adherens junctions at their apical borders and by gap junctions (valve-to-valve cell and valve to intestine) (PhaFIG 11).
PhaMOVIE 4: 3-D reconstruction of how VPI cells stack. Stacking of the six vpi cells from the anterior end (top left) toward the posterior is shown. Three-dimensional rendition of the cells based on tracings from serial TEMs. (Image source: B. Henick and A Hartley, based on MRC series.) Each cell is given a unique color. Colors do not follow the WormAtlas color code. Click on image to play movie.
No apparent muscular elements operate within the valve cells, nor are there any muscles attaching to this valve from the outside. Thus, the valve is probably a passively open and patent channel at all times, but rather narrow in caliber. The pm8 muscle of the pharynx is in appropriate position to act alone as a sphincter just rostral to the valve cells, but pm8 shows no direct innervation. Contraction of the pm8 has been suggested to open the valve wider when the grinder is active (Avery and Thomas, 1997). Hence, restrictions to regurgitation of ingested material may be due to operation of the pharyngeal grinder within the second bulb of the pharynx, immediately anterior to this valve. Squeezing motions of the terminal bulb would force food through the grinder and into the intestine via the pharyngeal-intestinal valve during the pumps when the pressure in terminal bulb is high. The intestinal lumen widens considerably just behind this region, which would result in a drop in internal pressure, which may limit any reflux from the intestine back through the valve.
Each valve cell has an electron-dense cytoplasm and occupies a thin wedge-shaped domain surrounding about one half of the lumen of the valve (PhaFIG 11). The nuclei of valve cells are flattened in shape, and the cytoplasm contains radial bands of intermediate filaments anchoring the apical cuticle to the basal lamina of the epithelium via hemidesmosomes, again in a very similar fashion to those seen in the buccal epithelium (D.H. Hall, unpublished).
9 Specific Structures Within the Pharynx
9.1 Channels of the Pharyngeal Lumen
These channels, three narrow grooves in outer corners of the pharyngeal cuticle, seem to allow an escape route for liquid to be regurgitated out of the pharynx via the buccal cavity. This results in food particles being trapped in the sieve while fluid is expelled. These channels extend under the mc1 marginal cells and connect into the buccal cavity (PhaFIG 2) (Albertson and Thomson, 1976).
9.2 Cuticle of the Pharynx
A thin cuticle lining extends to cover the interior surface of the pharyngeal passageway from the lips to the back of the pharynx, ending at the rear of the pharyngeal-intestinal valve. The pharyngeal cuticle is formed by the pharyngeal epithelium and the muscle cells to cover the apical surfaces of many cells acting in concert, much as the thickened pharyngeal basal lamina is formed jointly on basal surfaces of the pharyngeal cells. Unlike the body cuticle, this cuticle shows no layers; however, it shows some reinforcement at points of stress. For instance, it becomes keratinized to stain more densely by TEM at certain regions (PhaFIG 2). Other specialized portions of the pharyngeal cuticle include the following.
9.2.1 Bridging Cuticle
A small discrete region of cuticle connects the body wall cuticle covering the lips to the cuticle lining the buccal cavity. This bridging cuticle lies on the outer face of the anterior arcade and may also be touched briefly by the posterior arcade (see Epithelial System - Interfacial Cells; InterFIG 1).
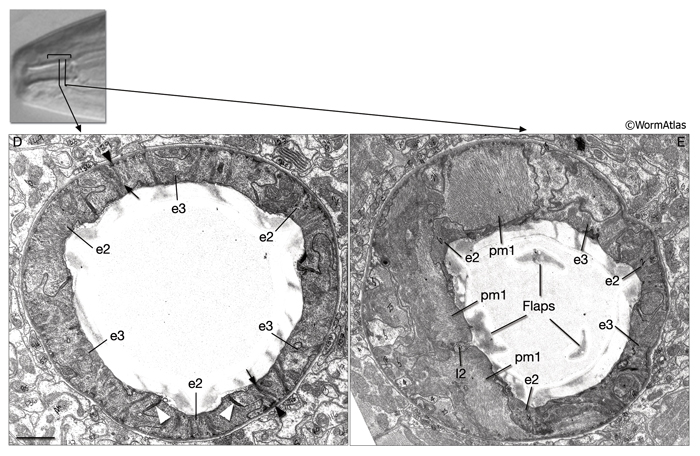
9.2.2 Flaps
These structures close the opening of the buccal cavity to the pharyngeal lumen and may correspond to the onchia or buccal teeth described in larger nematodes (see Chitwood and Chitwood, 1950; see also PhaFIG 2; PhaFIG 5; InterFIG 1). Three flaps extend inward from the level of from the pharyngeal muscle cells pm1 and pm2 to restrict flow at the rear of the buccal cavity. The flap cuticle is very electron dense, suggesting a sclerotic hardening to stiffen the flaps and the entryway to the true pharynx.
9.2.3 Grinder
The grinder is a cuticle specialization made primarily by pm6 and pm7 muscles. Its three contact zones, made by the three pairs of muscle cells, rotate when the muscles contract (Avery and Thomas, 1997). These interlocking "teeth" macerate the food and may function as a valve to regulate one-way traffic of food into the intestine (PhaFIG 2). Although the pm6 muscle filaments are oriented in radial fashion to the grinder, some portions of the pm7 muscles are oriented obliquely to the anterior-posterior axis and are anchored on the basal pole to the rear of the terminal bulb. These pull from the posterior side of the teeth, and coordinated action of these muscles may then rotate the grinder segments and force the teeth to scrape past and engage one another. The food caught and ground up between the teeth is passed back to the intestine through the pharyngeal-intestinal valve. Relaxation of the terminal bulb returns the grinder to its resting state.
9.2.4 Pharyngeal Sieve
These finger-like extensions from the pharyngeal cuticle probably act to trap bacteria (PhaFIG 2). They extend from the cell borders where pm4 muscles meet the neighboring mc1 cells. They project over a region of about 20 μm in length within the narrow lumen, ending near the transition of the metacorpus to the isthmus.
9.3 Gap Junctions
The muscle cells and the marginal cells of the pharynx are linked to and communicate with each other via elaborate gap junctions (Phelan, 2005; see also PhaFIG 7; PhaTABLE 1). Gap junctions also exist between the pharyngeal neurons and also between the pharyngeal neuron I1 and the extrapharyngeal neuron RIP. Gap junctions within the pharynx are composed of innexins, invertebrate gap junction proteins. Innexins heteromerically assemble into hexameric hemichannels and form pores between cells. This gap junction network confers a high level of connectivity within the pharynx, which is essential in coordinating waves of muscle contractions and spreading the neuronal input.
10 Feeding Behavior
C. elegans is a filter-feeder. Particles (bacteria) are taken in as suspended in liquid and then trapped in the pharynx while the liquid is expelled outside by the function of the corpus and anterior isthmus (Avery and Thomas, 1997; Avery and Shtonda, 2003). The particles are then transported to the terminal bulb, ground and passed into the lumen of the intestine. The feeding behavior consists of two motions; pumping, a contraction-relaxation cycle involving the corpus, anterior half of the isthmus and terminal bulb; and posterior isthmus peristalsis. Pumping involves near-simultaneous contraction of the muscles of the corpus, anterior isthmus and terminal bulb followed by near-simultaneous relaxation. When a feeding motion begins, contraction of the corpus and anterior isthmus opens their lumens, sucking particles and liquid in, whereas contraction of the terminal bulb muscles breaks up already-trapped bacteria and passes the debris posteriorly towards the intestine (Avery and Shtonda, 2003). At this stage corpus and anterior isthmus are separated hydrodynamically from the terminal bulb by a closed isthmus. During the relaxation period that follows, the lumens of the corpus and the anterior isthmus close, allowing for the liquid to be expelled through the radial channels while bacteria are retained and the grinder returns to its resting position. Unlike the corpus, which contracts and relaxes as a unit, the anterior isthmus contracts in a wave that proceeds rapidly from anterior to posterior. Also, the motions of the anterior isthmus are slightly delayed relative to the corpus, resulting in a net posterior transport of food particles during a pump (Avery and Shtonda, 2003). When the muscles again contract, the bacteria are carried further posteriorly by the inflow of liquid. Roughly one out of four pumps is followed by a posterior isthmus peristalsis where the trapped bacteria are carried from the anterior isthmus backwards to the grinder. Although each pm5 muscle cell runs the entire length of the isthmus, peristalsis occurs as a wave that propagates from anterior to posterior instead of simultaneously along its length. This capacity of isthmus for asynchronous contraction is suggested to permit the terminal bulb and corpus lumen to be at different pressures (Avery and Thomas, 1997).
Pumping is suppressed during the dauer larval stage and in response to a touch stimulus in adults. As described above, the latter response depends on an RIP/I1 connection. This circuit may also be important for suppression of pumping in the dauer larva (Keane and Avery, 2003). Inhibition of pumping during the dauer stage may save energy and prevent ingestion of environmental toxins when no appropriate food is available.
Under laboratory conditions, neural control of feeding can be managed completely by the pharyngeal nervous system. Three pharyngeal motor neuron types, M3, MC and M4, seem necessary and sufficient for normal feeding of the animal under these conditions. M3 neurons control the timing of the end of a pump, that is, initiation of pharyngeal relaxation. They are inhibitory-type motor neurons, and because of their free endings in the metacorpus, they are also suggested to be proprioceptive sensory neurons (Albertson and Thomson, 1976). Through a self-acting proprioceptive loop, M3 neurons are postulated to fire in response to corpus muscle contraction and cause muscle relaxation. M3 neurons are glutamatergic, and the fast inhibitory glutamatergic transmission from M3 neurons to the pharyngeal muscle is thought to be mediated through a glutamate-gated (ionotropic) chloride channel (Dent et al., 1997, 2000).
MC motor neurons control the rate of excitation of pharyngeal muscle. They mediate rapid pharyngeal pumping and are necessary for increased pumping in response to food (Avery and Horvitz, 1989). MC neurons are cholinergic excitatory motor neurons (Raizen et al., 1995; Keane and Avery, 2003). Mutants in which acetylcholine synthesis (cha-1) or packaging (unc-17) is disrupted show a pumping defect similar to that seen after MC ablation. Functional studies suggest that MC neurons synapse onto pm4 and that synaptic transmission from MC neurons to pm4 requires the nicotinic acetyl-choline receptor (nAChR) subunit, EAT-2 to be expressed on pm4. Mutations in eat-2 mimic MC cell loss (Keane and Avery, 2003; L. Avery, pers. comm.). MC neurons may also be mechanosensory, because they have free endings at the boundary between the procorpus and metacorpus and may sense the presence of bacteria in the pharynx.
The M4 neuron is required for posterior isthmus peristalsis. It synapses onto the posterior half of the isthmus muscles. M4 ablation leads to growth failure since the animals can not ingest any food (Avery and Horvitz, 1989). However, these animals continue to pump, and as a result the anterior isthmus and corpus become stuffed with bacteria. M4 is cholinergic but may also be utilizing another neurotransmitter since cholinergic antagonists and mutations that affect nicotinic ACh receptors only partially block M4 function (L. Avery, pers. comm.).
11 List of Pharyngeal Cells
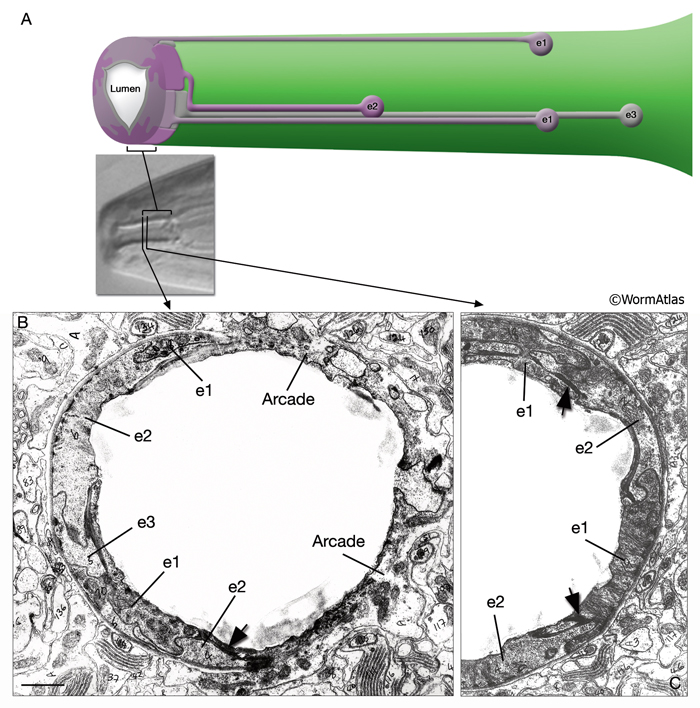
i. Buccal epithelium of the pharynx (e)
1. First epithelial ring
e1D
e1VL
e1VR
2. Second epithelial ring
e2DL
e2DR
e2V
3. Third epithelial ring
e3D
e3VL
e3VR
ii. Pharyngeal muscles (pm). Note that in earlier publications these cells are labeled as "m"; m1, m2, m3 etc., but here they are labeled "pm" for "pharyngeal muscle" (Avery and Thomas, 1997).
1. First pharyngeal muscle ring; all fuse into one syncytium around hatching
pm1DL
pm1DR
pm1L
pm1R
pm1VL
pm1VR
2. Second pharyngeal muscle ring
pm2DL; fuses with DR around hatching
pm2DR; fuses with DL around hatching
pm2L; fuses with VL around hatching
pm2R; fuses with VR around hatching
pm2VL; fuses with L around hatching
pm2VR; fuses with R around hatching
3. Third pharyngeal muscle ring
pm3DL; fuses with DR around hatching
pm3DR; fuses with DL around hatching
pm3L; fuses with VL around hatching
pm3R; fuses with VR around hatching
pm3VL; fuses with L around hatching
pm3VR; fuses with R around hatching
4. Fourth pharyngeal muscle ring
pm4DL; fuses with DR around hatching
pm4DR; fuses with DL around hatching
pm4L; fuses with VL around hatching
pm4R; fuses with VR around hatching
pm4VL; fuses with L around hatching
pm4VR; fuses with R around hatching
5.Fifth pharyngeal muscle ring
pm5DL; fuses with DR around hatching
pm5DR; fuses with DL around hatching
pm5L; fuses with VL around hatching
pm5R; fuses with VR around hatching
pm5VL; fuses with L around hatching
pm5VR; fuses with R around hatching
6. Sixth pharyngeal muscle ring
pm6D
pm6VL
pm6VR
7. Seventh pharyngeal muscle ring
pm7D
pm7VL
pm7VR
8. Eighth pharyngeal muscle ring
pm8
iii. Marginal cells (mc)
1. First marginal cell ring
mc1DL
mc1DR
mc1V
2. Second marginal cell ring
mc2DL
mc2DR
mc2V
3. Third marginal cell ring (syncytial, cells fuse around hatching)
mc3DL
mc3DR
mc3V
iv. Pharyngeal-intestinal valve (vpi)
1. First pharyngeal valve ring
vpi1
2. Second pharyngeal valve ring
vpi2DL
vpi2DR
vpi2V
3. Third pharyngeal valve ring
vpi3D
vpi3V
v. Pharyngeal neurons
1. Motor neurons
M1
M2L/R
M3L/R
M4
M5
2. Interneurons
I1L/R
I2L/R
I3
I4
I5
I6
2. Other neurons
MI
NSML/R
MCL/R
vi. Pharyngeal glands (g)
1. First pharyngeal gland ring
g1AL; ventral left g1 gland cell
g1AR; ventral right g1 gland cell
g1P; dorsal gland cell
2. Second pharyngeal gland ring
g2L
g2R
See also PharynxAtlas
12 References
Albertson, D.G. and Thomson, J.N. 1976. The pharynx of Caenorhabditis elegans. Phil. Trans. Royal Soc. London 275B: 299-325. Article
Avery D.G. and Thomas, J.H. 1997. Feeding and defecation. In C. elegans Volume II. Ed.s Riddle D.L., Blumenthal, T., Meyer B.J. and Priess J.R . Pp 679-716. Cold Spring Harbor Laboratory Press. Article
Avery, L. and Horvitz, H R. 1987. A cell that dies during wild-type C. elegans development can function as a neuron in a ced-3 mutant. Cell. 51: 1071-1078. Abstract
Avery, L. and Horvitz, H.R. 1989. Pharyngeal pumping continues after laser killing of the pharyngeal nervous system of C. elegans. Neuron 3: 473-485. Abstract
Avery, L. and Shtonda, B.B. 2003. Food transport in the C. elegans pharynx. J. Exp. Biol. 206: 2441-2457. Article
Avery, L. Bargmann, C.I. and Horvitz, H.R. 1993. The Caenorhabditis elegans unc-31 gene affects multiple nervous system-controlled functions. Genetics 134: 454-464. Article
Bird, A.F. and Bird, J. 1991 The structure of nematodes. Academic Press, San Diego, CA.
Bucher, E.A. and Seydoux, G.C. 1994. Gastrulation in the nematode Caenorhabditis elegans. Seminars in Developmental Biology. 5: 121-130. Abstract
Chalfie, M., Sulston, J.E., White, J.G., Southgate, E., Thomson, J.N. and Brenner, S. 1985. The neural circuit for touch sensitivity in Caenorhabditis elegans. J. Neurosci. 5: 956-964. Article
Chitwood, B.G. and Chitwood, M.B. 1950. An introduction to nematology. Baltimore, University Park Press.
Dent, J.A., Davis, M.W. and Avery, L. 1997. avr-15 encodes a chloride channel subunit that mediates inhibitory glutamatergic neurotransmission and ivermectin sensitivity in Caenorhabditis elegans. EMBO Journal 16: 5867-5879. Article
Dent, J.A., Smith, M.M., Vassilatis, D.K. and Avery, L. 2000. The genetics of ivermectin resistance in Caenorhabditis elegans. Proc. Natl. Acad. Sci. USA 97: 2674-2679. Article
Franks, C.J., Holden-Dye, L., Bull, K., Luedtke, S. and Walker, R.J. 2006. Anatomy, physiology and pharmacology of Caenorhabditis elegans pharynx: a model to define gene function in a simple neural system. Invert. Neurosci. 6: 105-122. Abstract
Hall, D.H. and Hedgecock, E.M. 1991. Kinesin-related gene unc-104 is required for axonal transport of synaptic vesicles in C. elegans. Cell 65: 837-847. Abstract
Hedgecock, E.M. and Thomson, J.N. 1982. A gene required for nuclear and mitochondrial attachment in the nematode C. elegans. Cell. 30: 321-330. Abstract
Horner, M.A., Quintin, S., Domeier M.E., Kimble, J.E., Labouesse, M. and Mango, S.E. 1998. pha-4, an HNF-3 homolog, specifies pharyngeal organ identity in Caenorhabditis elegans. Genes Dev. 12: 1947-1952. Article
Keane, J. and Avery, L. 2003. Mechanosensory inputs influence Caenorhabditis elegans pharyngeal activity via ivermectin sensitivity genes. Genetics 164: 153-162. Article
Knight, C.G., Patel, M.N., Azevedo, R.B.R. and Leroi, A.M. 2002. A novel mode of ecdysozoan growth in Caenorhabditis. Evol. Dev. 4: 16-27. Abstract
Koppen, M., Simske, J.S., Sims, P.A., Firestein, B.L., Hall, D.H., Radice, A.D., Rongo, C. and Hardin, J.D. 2001. Cooperative regulation of AJM-1 controls junctional integrity in Caenorhabditis elegans epithelia. Nature Cell Biol. 3: 983-991. Abstract
Leung, B., Hermann, G.J, Priess, J.R. 1999. Organogenesis of the Caenorhabditis elegans intestine. Dev. Biol. 216: 114-134. Article
Mango, S.E. 2007. The C. elegans pharynx: a model for organogenesis. WormBook, ed. The C. elegans Research Community, WormBook, doi/10.1895/wormbook.1.129.1. Article
Phelan, P. 2005. Innexins: members of an evolutionarily conserved family of gap-junction proteins. Biochim. Biophys. Acta 1711: 225-245. Abstract
Portereiko, M.F. and Mango, S.E. 2001. Early morphogenesis of the Caenorhabditis elegans pharynx. Dev. Biol. 233: 482-494. Article
Raizen, D.M., Lee, R.Y.N. and Avery, L. 1995. Interacting genes required for pharyngeal excitation by motor neuron MC in Caenorhabditis elegans. Genetics 141: 1365-1382. Article
Smit, R. B., Schnabel, R. and Gaudet, J. 2008. The HLH-6 transcription factor regulates C. elegans pharyngeal gland development and function. PLoS Genetics 4(10): e1000222. Article
Sulston, J.E., Schierenberg, E., White, J.G. and Thomson, J.N. 1983. The embryonic cell lineage of the nematode Caenorhabditis elegans. Dev. Biol. 100: 64-119. Article
Wright, K.A. and Thomson, J.N. 1981. The buccal capsule of C. elegans (Nematoda: Rhabditoidea): An ultrastructural study. Can. J. Zool. 59: 1952-1961. Article
|

 Click pictures for new window with figure and legend, click again for high resolution image
Click pictures for new window with figure and legend, click again for high resolution image