|
|
|
THE DAUER EXCRETORY SYSTEM
 Click pictures for new window with figure and legend, click again for high resolution image Click pictures for new window with figure and legend, click again for high resolution image
1 Overview
The C. elegans excretory system carries out a number of important functions. First, the excretory system provides a means for concentrating and expelling metabolic wastes. Second, expulsion of excess water enables regulation of internal osmolarity, suggesting the excretory system functions similarly to the mammalian kidney. Third, exsheathment fluid, used for releasing the molted cuticle during molts, can be expelled via the excretory system. Finally, the excretory system secretes some hormones for export to target tissues (Nelson et al., 1983).
2 Anatomy
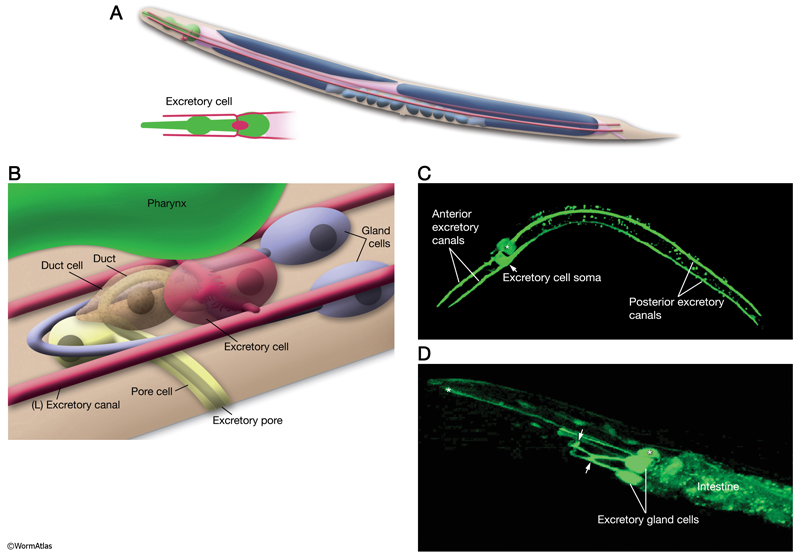
Four cell types make up the C. elegans excretory system. These are (1) the large, H-shaped excretory canal cell, which extends canals bilaterally from the animal’s head to tail, (2) the pulsatile excretory duct cell, (3) the pore cell adjacent to the body surface and (4) two fused gland cells (DExcFIG 1) (see also Hermaphrodite Excretory System) (Nelson et al., 1983). A specialized adherens junction is formed between the gland, duct and excretory cells, called the secretory-excretory junction or the secretory sinus. This structure permits the one-way flow of materials from the canal cell and gland cells into the duct and then out of the body through the excretory pore.
DExcFIG 1: Overview of C. elegans excretory system. A. Schematic showing the full length of the H-shaped excretory cell as seen from lateral side. In an adult animal, the excretory canals reach from the nose of the nematode to the tail region. (Inset) Head portion from the ventral side. B. The excretory system (lateral oblique view) consists of the fused pair of gland cells (blue), the excretory (canal) cell (red), the duct cell (brown) and the pore cell (yellow). Somata of all of these cells are located in the head region. The excretory cell is the largest cell in the nematode and is located juxtaposed to the terminal bulb of the pharynx on the ventral side. C. Epifluorescent image of a transgenic adult animal (dorsal oblique view) expressing the F22E10.1::GFP reporter in the excretory cell. Asterisk marks the pharyngeal gland, which also expresses GFP. (Image source: R. Newbury and D. Moerman.) D. Epifluorescent micrograph of an animal showing expression of B0403.4::GFP in the excretory gland cell, ventral oblique view. The two small arrows point to regions where two cells fuse to make one syncytial cell. The anterior arrow marks the region where the excretory gland cell is suggested to receive synaptic input from the nerve ring. Pharyngeal gland cells also express this marker (asterisks). (Image source: R. Newbury and D. Moerman.)
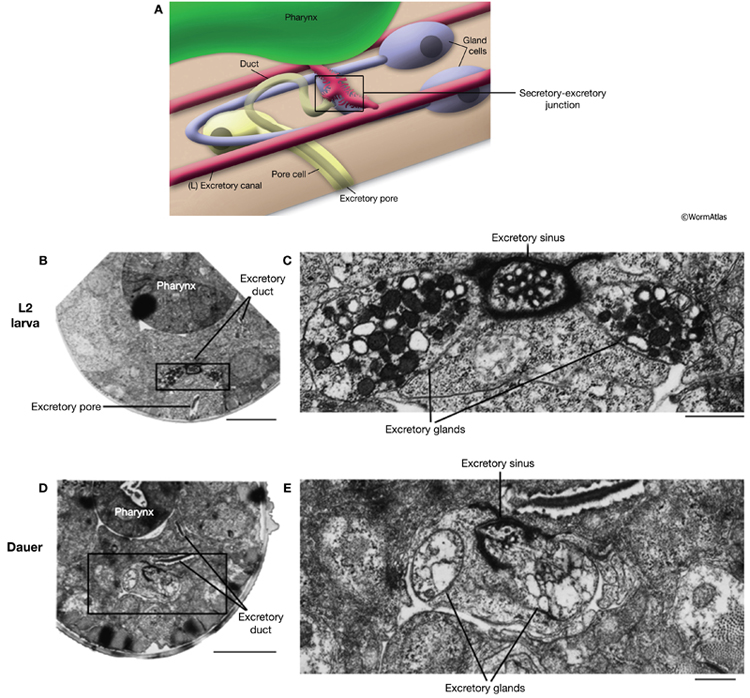
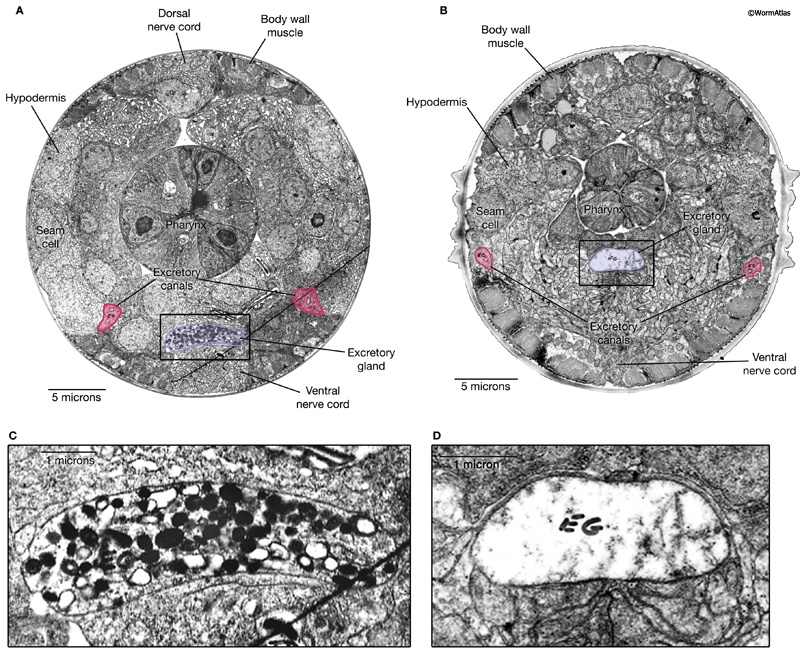
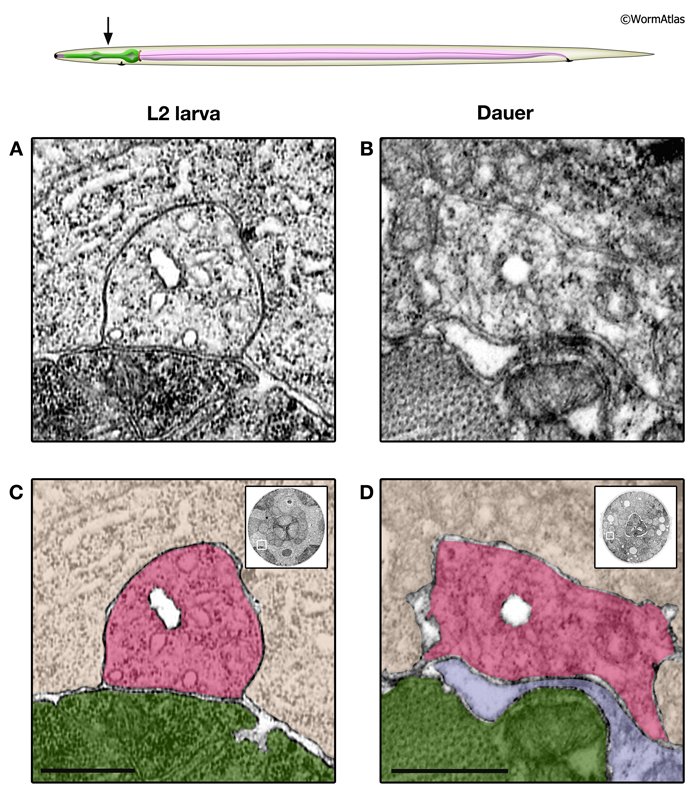
Overall, the cells of the excretory system retain the same positions and structures in dauer and non-dauer stages. However, there are subtle differences in the form and function of the dauer excretory system. In developing larvae and adults, the gland cell contains numerous secretory granules, which are positioned adjacent to the secretory-excretory junction. In dauers, the gland cell cytoplasm is devoid of secretory granules. Instead, the gland cell contains loosely arranged membranes which may be remnants of the granule-containing state (DExcFIG 2) (Nelson et al., 1983; Albert and Riddle, 1988). Excretory gland granules are also missing from the excretory glands in the area of the secretory-excretory junction and sinus (DExcFIG 3). The absence of secretory granules in dauer gland cells is unlikely to be due to starvation, because granules were still present in the gland cell of a starved L2. Rather, the loss of granules appears to be a dauer stage-specific characteristic.
DExcFIG 3: Secretory-excretory junction in L2 and dauer larvae. A. Illustration showing the overall structure of the secretory-excretory junction in the head near the pharynx. The boxed region indicates the location of the secretory sinus and is the approximate region shown in panels B-E (below). B&D. Transverse sections of the head region where the secretory sinus is located (boxed region) in an L2 (B) and dauer larva (D). The excretory pore is visible in the L2 section (B) and is slightly offset in the dauer section (D). Bars, 5 microns. C&E. Close-up views of the boxed regions from B&D showing the secretory sinus and adjacent excretory gland cells. Dark-staining granules are present in the excretory gland cells of the L2 (C) but absent in the dauer (E). Bars, 1 micron. (Image sources: L2 [D. Riddle] L2 28-14 #1315; dauer [D. Riddle] 50-2-2 #1049.)
In well-fed, developing larvae and adults, the excretory gland cells are strongly stained by PAF (paraldehyde-fuchsin), particularly in the region where secretory granules are located (Nelson et al., 1983; for detailed images see FIGs 19-24 in Nelson et al., 1983). This staining is affected by food-deprivation in dauer and non-dauer larvae, as PAF staining of the excretory gland is diminished or absent in larvae that were starved for 6 hours or more and in dauers. |
3 Function
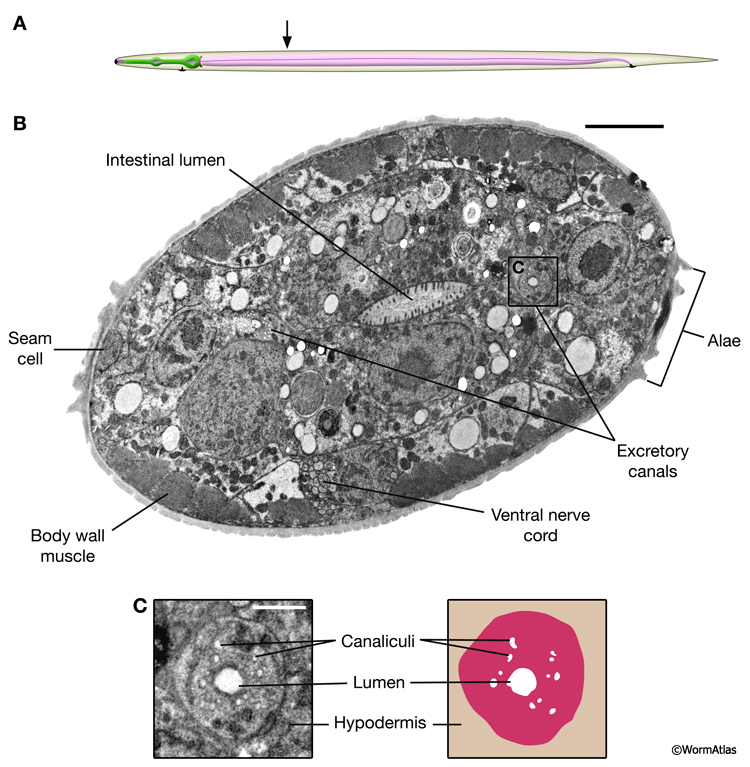
Several lines of evidence indicate that the excretory system is still active in dauers. First, the excretory pore in dauers is open to the outside environment allowing materials to transit out of the animal in this stage (Cox et al., 1981; see section on Dauer Cuticle of Excretory Pore). This contrasts to closure of the buccal opening by the thick dauer cuticle. Second, the dauer excretory duct is observed to pulsate, and this pulsation is more prominent in dauers than in other stages (Nelson et al., 1983; Nelson and Riddle, 1984; Holt and Riddle, 2003). The excretory system may excrete water from the dauer body in order to maintain the high specific density characteristic of this stage (Nelson et al., 1983). Duct pulsation is more active in dauers placed in a medium with low osmolarity, such as distilled water, than higher osmolarity, such as M9 buffer (Nelson and Riddle, 1984). The excretory canals retain the circular lumen and plentiful canaliculi, also suggestive of active function in this stage (DExcFIG 4 and DExcFIG 5).
DExcFIG 5: Excretory canal structure in a dauer larva. Cross-section view from a dauer larvae in the midbody region highlighting bilaterally symmetric excretory canals. A. Illustration showing approximate position of section within the dauer body. B. Transverse, slightly oblique, section showing the midbody region of a dauer larva. In the midbody, bilateral excretory canals are positioned adjacent to the seam cells, which are easily identified by the dauer cuticular alae. Positions of the intestinal lumen, body wall muscle and ventral nerve cord are also identified. Scale bar, 5 microns. C. Left panel shows an enlarged view of the excretory canal in cross-section, in which the circular lumen and canaliculi of the canal cell are visible. The right panel shows a graphical representation of the canal cell and internal components. Scale bar, one micron. (Image source: [D. Hall] him-5dar_1127-1_74535_B5_029.)
|
The excretory gland is dispensable for dauer formation and recovery. Laser ablation of the excretory gland did not alter dauer formation under dauer-inducing conditions nor in dauer-constitutive genetic mutants (Nelson and Riddle, 1984). Furthermore, ablation of the gland cells did not impair recovery from the dauer stage (Nelson and Riddle, 1984). These experiments, however, do not rule out the possibility that the excretory system participates in the production of the secreted dauer pheromone which functions to communicate developmental decisions to other members of the population (Nelson et al., 1983). More recent study of the dauer pheromone system reveals that ascarosides are the active component of dauer pheromone (see section on Dauer Pheromone). Expression analysis of the ascaroside biosynthetic pathway indicates that multiple tissues produce these pheromones, including the intestine, hypodermis and possibly muscle (Butcher et al., 2009). The excretion pathway for these molecules has not been traced, but could involve excretion through the alimentary system at the rectum or through the excretory system at the excretory duct.
4 References
Albert, P.S. and Riddle, D.L. 1983. Developmental alterations in sensory neuroanatomy of the Caenorhabditis elegans dauer larva. J. Comp. Neurol. 219:461-481. Abstract
Butcher, R.A., Ragains, J.R., Li, W., Ruvkun, G., Clardy, J. and Mac, H.Y. 2009. Biosynthesis of the Caenorhabditis elegans dauer pheromone. Proc. Natl. Acad. Sci. 106: 1875-1879. Article
Cox, G.N., Staprans, S. and Edgar, R.S. 1981. The cuticle of Caenorhabditis elegans. II. Stage-specific changes in ultrastructure and protein composition during postembryonic development. Dev. Biol. 86: 456-470. Article
Holt, S.J. and Riddle, D.L. 2003. SAGE surveys C. elegans carbohydrate metabolism: evidence for an anaerobic shift in the long-lived dauer. Mech. Age. Develop. 124: 779-800. Abstract
Nelson, F.K. and Riddle, D.L. 1984. Functional study of the Caenorhabditis elegans secretory-excretory system using laser microsurgery. J. Exp. Zool. 231: 45-56. Abstract
Nelson, F.K., Albert, P.S. and Riddle, D.L. 1983. Fine structure of the Caenorhabditis elegans secretory-excretory system. J. Ultrastruct. Res. 82: 156-171. Article
|

This chapter should be cited as: Wolkow, C.A. and Hall, D.H. 2014. The Dauer Excretory System. In WormAtlas. doi:10.3908/wormatlas.3.8
Edited for the web by Laura A. Herndon. Last revision: April 29, 2014. |

|
|
|
|