|
|
|
THE AGING CUTICLE
 Click pictures for new window with figure and legend, click again for high resolution image Click pictures for new window with figure and legend, click again for high resolution image
1 General Description
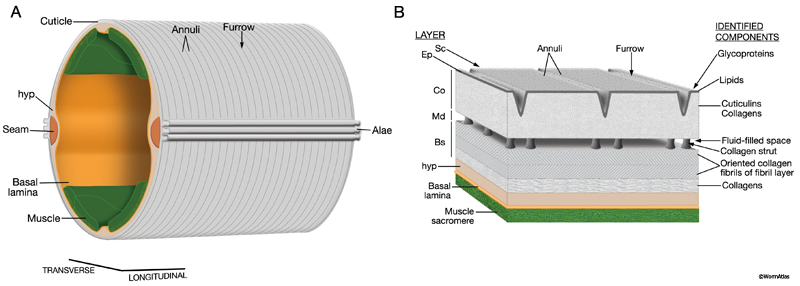
The C. elegans cuticle protects the animal from environmental threats and provides support to the body. Around the body, the cuticle forms small circumferential furrows separated by broader ridges, called annuli. This arrangement is interrupted in particular areas by pores and swellings that allow specialized neurons to access the environment. Major body openings to the environment are lined by cuticle. These include the buccal opening at the head where nutrients enter the body, the excretory pore, the opening of the vulva where eggs are laid and the rectum in the tail. Over most of the length of the adult body, the cuticle features linear alae on the left and right sides, in register with the seam cells. Since the animal always lies on one side or the other, one set of alae are always facing downward towards the substrate, and are presumed to provide the adult added traction during body motion over solid surfaces. (For more background, please refer to WormAtlas Hermaphrodite section on Cuticle.)
Over most of the body, the components of the cuticle are secreted by the hypodermis and seam cells, which are the epithelial cells covering the body (ACutFig 1A). (For details, see Hypodermis and Seam Cells). In young adults, the body cuticle is approximately 0.5 microns thick and is composed of five zones (ACutFig 1B) (Cox et al., 1981). The outermost layer is the thin surface coat containing glycoproteins. Next is the epicuticle layer, which is a membrane layer containing lipids and glycolipids. Below the epicuticle layer lies the cortical zone, made up of cuticulins and collagens. The medial zone is a fluid-filled space with periodic collagenous struts that help separate and secure the cortical and basal zones with respect to one another (Kramer, 1997). The innermost layer of the cuticle is the basal zone which, itself, consists of three regions. Two exterior fibrous layers contain oriented collagen fibrils that overlie a layer of collagen above the hypodermis. The cuticle that covers the major body openings, such as the anus, excretory pore, vulva and pharynx, is produced by interfacial cells that line these structures. While this type of cuticle does not appear to be composed of layers, it still provides adequate structural support for function. Overall, the young adult cuticle appears to be both tough yet flexible, resisting outside insults that might tear it open or damage internal organs and insulating the animal from noxious agents. The cuticle also serves as the animals exoskeleton, giving more rigidity than any internal structure, and perhaps balancing local forces during muscle contractions (Kramer, 1997). The body muscles themselves are firmly secured to the overlying cuticle via several forms of periodic attachment sites (see CutFIG 10).
|
2 Changes in the Cuticle with Age
Adult animals do not molt, so the adult cuticle persists throughout the entire adult lifespan. Across the body, the cuticle continues to grow during adulthood and into older age (Herndon et al., 2002; Herndon et al., 2017). Continued growth causes the cuticle to thicken and cuticle growth may continue until the hypodermis and seam are no longer capable of secreting cuticular components. This continuous growth may result from unregulated biosynthesis of cuticle-related proteins as post-reproductive shut down of overall expression does not appear to occur (Herndon et al., 2002). The cuticle in older animals also appears wrinkled, possibly due to continuous growth and/ or to shrinkage of internal body tissues during aging (ACutFIG 2A-D; Herndon et al., 2002; Herndon et al., 2017). Cuticle wrinkles may arise from the combined effects of a weaker, thinner hypodermis, loosened connections between the cuticle and hypodermis, and weakening muscles.
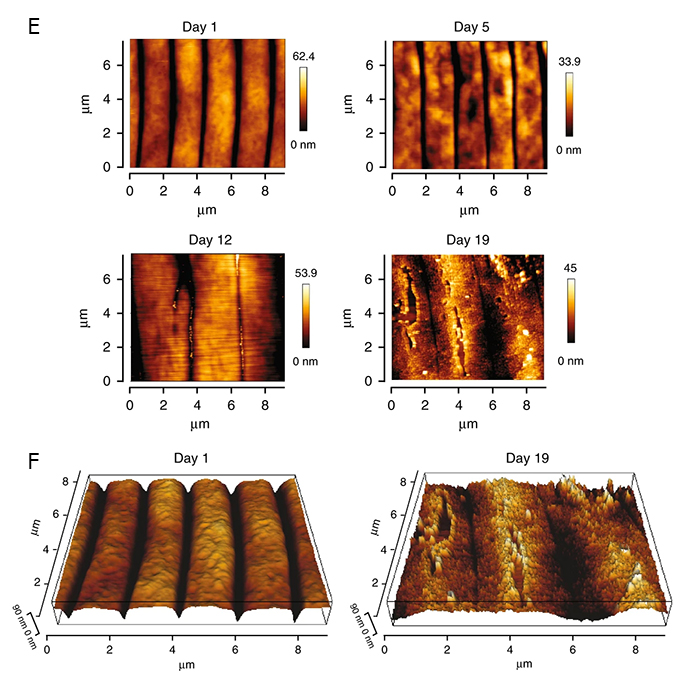
Additionally, studies using Atomic Force Microscopy (AFM) have quantitatively measured the changes in biomechanical properties of the cuticle in young and old C. elegans (Essmann et al., 2020) and found that the cuticle loses its stiffness and integrity with age. Topographical images reveal that the cuticle becomes rougher and develops breaks and cavities along its surface with an accompanying flattening of the annular furrow structure (ACutFIG 2E&F).
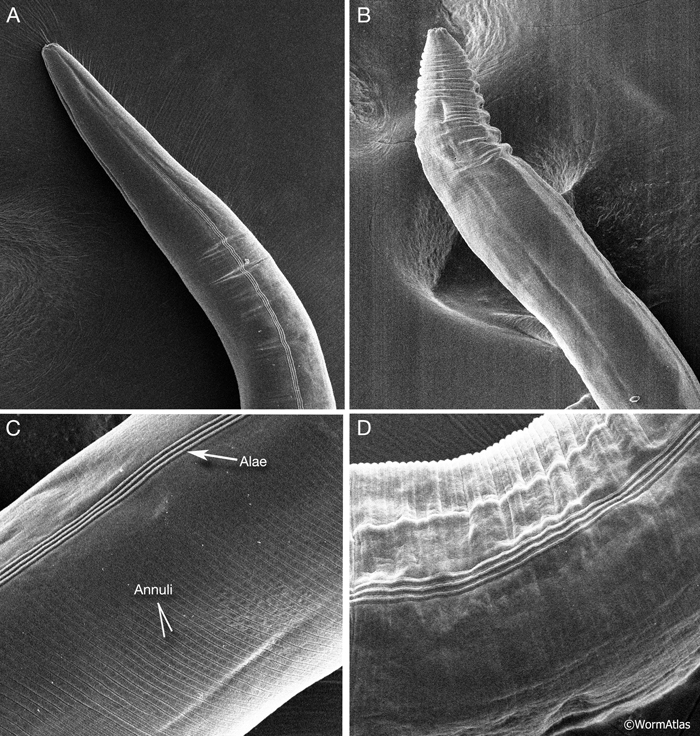
ACutFIG 2A-D: Scanning electron micrographs (SEMs) of young and old C. elegans showing increased wrinkling in the cuticle. A&C. Head and body regions, respectively, of young adult animals. B&D. Head and body regions of older adults showing wrinkled appearance of the cuticle (cf. Hall et al., 1999). The image of the cuticle from an older animal shown here reveals minimal to modest wrinkling, but often one finds local regions with much deeper wrinkles, regardless of fixation protocol or whether viewed by SEM or TEM, suggesting that deep wrinkling is not a technical artifact. (Image source: S. Melov.)
ACutFIG 2E&F: Atomic Force Microscopy (AFM) images of young and old C. elegans. E. AFM cuticle topography images of C. elegans cuticle at day 1, 5, 12 and 19 of adulthood. Topographical images show a decline in cuticle integrity with age as indicated by an increase in roughness with breaks and cavities appearing in older animals. F. 3-D topographical images show a flattening of the annuli furrow structure when old, 19 day C. elegans are compared to young, 1 day animals (Image source: Essmann et al., 2020; figure is modified from published version Nat Commun 2020 11, 1043 https://doi.org/10.1038/s41467-020-14785-0.) |
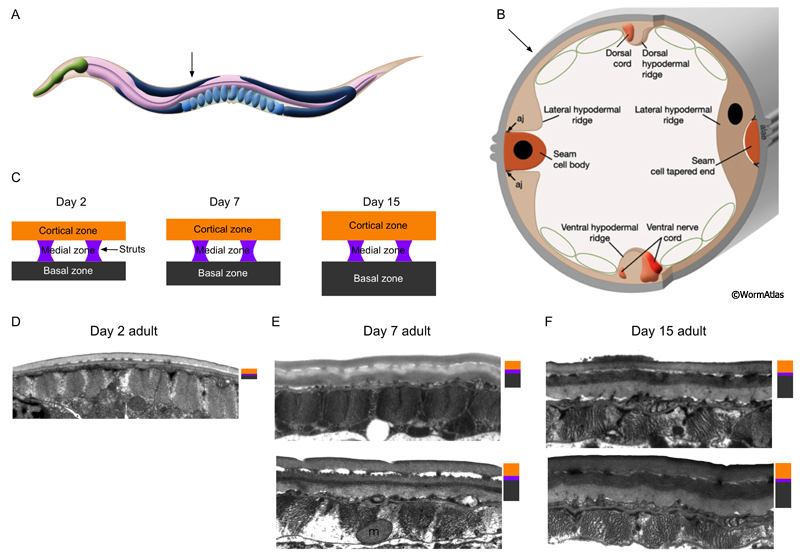
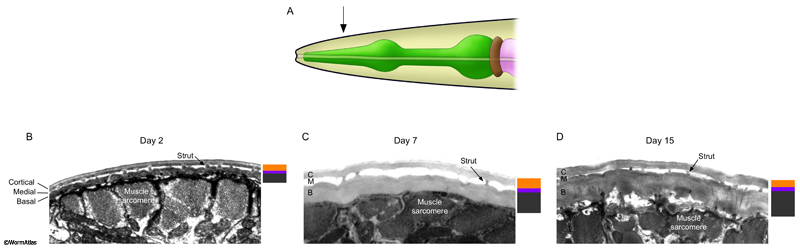
When viewed in cross section, all of the cuticle layers appear intact in the older adult. The most significant thickening is usually observed in the innermost basal zone, adjacent to the hypodermis (ACutFIG 3 and ACutFIG 4). Growth also continues in the outer cortical zone, although less consistently. The medial zone is relatively stable in appearance through adulthood, although the struts may become more irregularly spaced. Changes have not been observed in the morphology of the surface coat or epicuticle layers although this does not exclude the possibility of changes to the molecular components of these layers.
ACutFIG 3: Cuticle growth in the midbody region during aging. The cuticles cortical and basal zones continue to grow through adulthood. A. Adult hermaphrodite with arrow indicating approximate position of midbody sections shown in (D, E and F). B. Diagram of cross-section through the hermaphrodite midbody with arrow showing approximate position of sections shown in (D, E and F). C. Diagram showing expansion of the cuticles cortical (orange) and basal (black) zones between days 2 and 15 of adulthood. D, E and F. Transverse TEM sections of the cuticle overlying the body wall muscle in the midbody region in young (day 2) adults (D), middle-aged (day 7) adults (E) and older (day 15) adults (F). The sizes of the cuticle zones are shown beside each image; cortical (orange), medial (purple), basal (black). aj, adherens junction; m, muscle cell mitochondrion. (Image sources: D, N506 [D. Hall] N278; E, upper N825 [D. Hall] C0395, class A; lower N821 [D. Hall] 4994, class B; F, upper N816 [D. Hall] H0012, class B; lower H0016.) Images were scaled relative to the body muscle myofilaments which remain stable during adulthood.
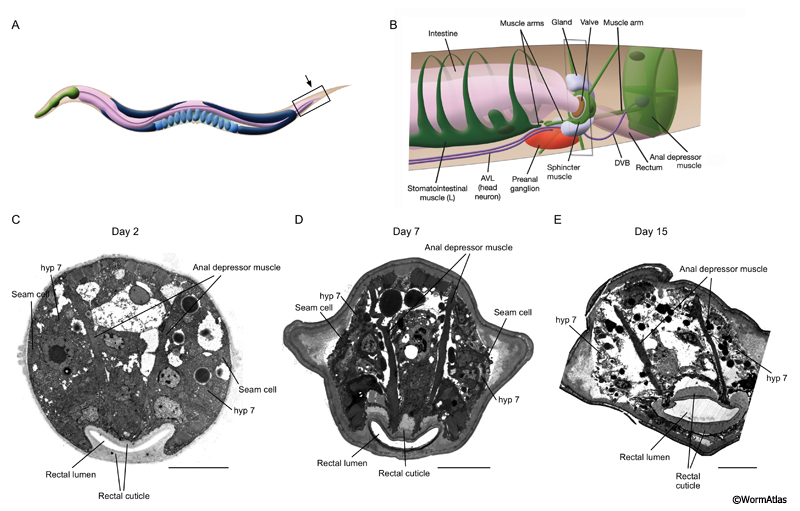
 Growth during later adulthood causes the basal zones to appear less orderly. The outer fibril layers become more loosely structured. The collagen layer adjacent to the hypodermis develops swellings in areas adjacent to the hypodermis, which could represent cuticle expansion into voids in the body surface. In older animals, the distinct cuticle linings of body openings remain relatively intact, and are possibly reinforced and expanded via the thickening process. This is especially evident in the rectum where the rectal cuticle may extend to more fully surround the anal depressor muscles (ACutFIG 5). Images of the tail tip in ACutFIG 5 help to emphasize the gradual loss of integrity of virtually all internal tissues, including the hypodermis and the anal depressor muscle sarcomeres, while the rectal cuticle remains strong and thus a barrier to entry by live bacteria from the outside environment.
One consequence of thickened body cuticle to older adults could be to limit motility. Movement rates decline with aging (Croll et al., 1977; Duhon and Johnson, 1987; Herndon et al., 2002; Huang et al., 2004). Aging-related locomotory decline has been attributed to muscle deterioration (Herndon et al., 2002; Glenn et al., 2004) and to synaptic changes (Toth et al., 2012; Morsci et al., 2016), yet stronger resistance from a thickened, inflexible cuticle could also negatively impact motility. Thickening of the rectal cuticle could similarly impact defecation behavior, which has been shown to decline with age (Croll et al., 1977; Bolanowski et al., 1981; Felkai et al., 1999).
ACutFIG 5: Rectal cuticle growth during aging. During adulthood, the rectal cuticle may expand into the body space to partially encompass the anal depressor muscle attachments. A. Adult hermaphrodite with rectal area indicated by box. Black arrow indicates approximate position of transverse TEM sections shown in C, D & E. B. Detail of the rectal area from boxed region in A. (Image source: WormAtlas). C, D & E. Transverse TEM sections of rectum where rectal cuticle joins the body cuticle at the posterior. Rectal cuticle is indicated. In older adults, the rectal cuticle expands into the body surrounding the base of the anal depressor muscles. Scale bars, 5 microns. (Image sources: C. B140C [D. Hall] T565; D. N826 [D. Hall] G5802, class A; E. N801 [D. Hall] E556, class B.) |
3 References
Blaxter, M.L. and Robertson W.M. 1998. The Cuticle. In The Physiology and Biochemistry of Free-living and Plant-parasitic Nematodes, (ed. R.N. Perry and D.J. Wright) pp. 25-48. CAB International, New York. Abstract
Bolanowski, M.A., Russell, R.L. and Jacobson, L.A. 1981. Quantitative measures of aging in the nematode Caenorhabditis elegans. I. Population and longitudinal studies of two behavioral parameters. Mech. Ageing Dev. 15: 279–295. Abstract
Cox, G.N., Kusch, M. and Edgar, R.S. 1981a. Cuticle of C. elegans: Its isolation and partial characterization. J. Cell Biol. 90: 7-17. Article
Croll, N.A., Smith, J.M. and Zuckerman, B.M. 1977. The aging process of the nematode Caenorhabditis elegans in bacterial and axenic culture. Exp. Aging Res. 3: 175–189. Abstract
Duhon, S.A. and Johnson, T.E. 1995. Movement as an index of vitality: comparing wild type and the age-1 mutant of Caenorhabditis elegans. J. Gerontol. Biol. Sci. 50:B254–B261. Abstract
Essmann C.L., Martinez-Martinez, D., Pryor, R., Leung, K.Y., Krishnan, K.B., Lui, P.P., Greene, N.D.E., Brown, A.E.X., Pawar, V.M., Srinivasan, M.A. and Cabreiro, F. 2020. Mechanical properties measured by atomic force microscopy define health biomarkers in ageing C. elegans. Nature Comm. 11: 1043. doi: 10.1038/s41467-020-14785-0. Article
Felkai, S., Ewbank, J.J., Lemieux, J., Labbe, J.C., Brown, G.G. and Hekimi, S. 1999. CLK-1 controls respiration, behavior and aging in the nematode Caenorhabditis elegans. EMBO J. 18: 1783–1792. Article
Glenn, C.F., Chow, D.K., David, L., Cooke, C.A., Gami, M.S., Iser, W.B., Hanselman, K.B., Goldberg, I.G. and Wolkow, C.A. 2004. Behavioral deficits during early stages of aging in Caenorhabditis elegans result from locomotory deficits possibly linked to muscle frailty. J. Gerontol. 59A: 1251-60. Article
Herndon, L.A., Schmeissner, P.J., Dudaronek, J.M., Brown, P.A., Listner, K.M., Sakano, Y., Paupard, M.C., Hall, D.H. and Driscoll, M. 2002. Stochastic and genetic factors influence tissue-specific decline in ageing C. elegans. Nature 419: 808-814. Article
Herndon, L.A., Wolkow, C.A., Driscoll, M. and Hall, D.H. 2017. Effects of ageing on the basic biology and anatomy of C. elegans. In Ageing: lessions from C. elegans. (ed Olsen, A. and Gill, M.). Chapter 2. pp. 9-39. Springer International, Switzerland. Abstract
Huang, C., Xiong, C. and Kornfeld, K. 2004. Measurements of age-related changes of physiological processes that predict lifespan of Caenorhabditis elegans. Proc. Natl. Acad. Sci. U S A. 101:8084–8089. Article
Kramer, J.M. 1997. Extracellular matrix. In C. elegans II (ed. D.L Riddle et al). pp. 471-500. Cold Spring Harbor Laboratory Press, Cold Spring Harbor, New York. Article
Lints, R. and Hall, D.H. 2009. The cuticle. In WormAtlas. doi:10.3908/wormatlas.1.12
Morsci, N.S., Hall, D.H., Driscoll, M. and Sheng, Z.H. (2016) Age-related phasic patterns of mitochondrial maintenance in adult Caenorhabditis elegans neurons. J. Neurosci. 36:1373-85. doi: 10.1523/JNEUROSCI.2799-15.2016 Article
Toth, M.L., Melentijevic, I., Shah, L., Bhatia, A., Lu, K., Talwar, A., Naji, H., Ibanez-Ventoso, C., Ghose, P., Jevince, A., Xue, J., Herndon, L.A., Bhanot, G., Rongo, C., Hall, D.H. and Driscoll, M. 2012. Neurite sprouting and synapse deterioration in the aging Caenorhabditis elegansnervous system. J. Neurosci. 32: 8778-90. Article
* Description of Behavioral Classes (A, B, C) as described in Herndon et al., 2002
To characterize aging phenotypes, age-synchronized individual worms were scored both for spontaneous movement and for response to prodding with a wire over the course of their lifespan. Three distinct classes representing behavioral phenotypes were established. Animals that move constantly and make sinusoidal tracks were designated as class A. Class B animals mainly move when prodded. When they move it is with uncoordinated motion, leaving non-sinusoidal tracks. Class C animals do not move forward or backward, even upon prodding, but do show head and/or tail movement and twitch in response to touch. All animals begin adulthood in class A. Class B animals appear around days 6-7 of adulthood and class C around day 9-10 (at 20oC). At later ages, animals representing all classes can be found within the same population and it was found that the behavioral class type was the better predictor of life expectancy than chronological age (Herndon et al., 2002). Due to the stochastic nature of the aging process in an individual nematode, these classifications only reflect ongoing changes in nerve and muscle, while other tissues can show very different age-related effects within one behavioral class, declining faster or remaining healthy much longer.
|

This chapter should be cited as: Wolkow, C.A, Herndon, L.A. and Hall, D.H. 2017. The aging cuticle. In WormAtlas. doi:10.3908/wormatlas.8.2
Edited for the web by Laura A. Herndon. Last revision: June 25, 2020. |

|
|
|
|